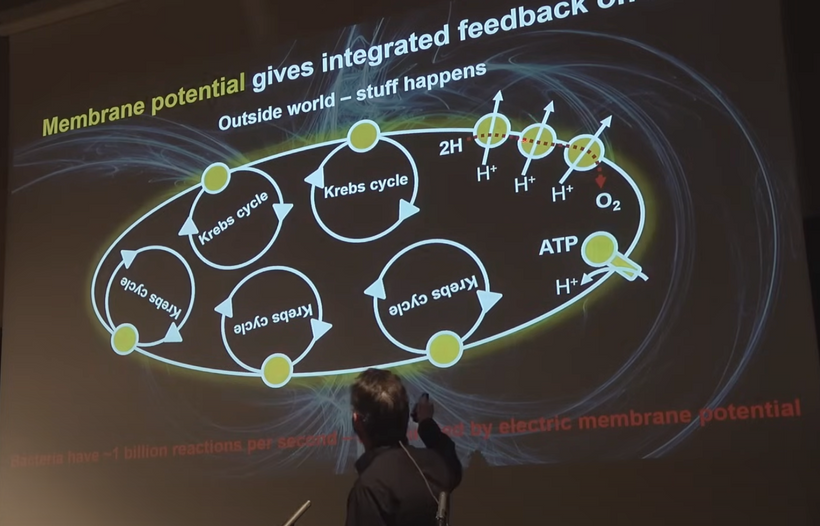
My decentralized synthesis powerfully reframes cancer, and ALL chronic disease broadly, as a failure of circadian-light entrainment leading to lost metabolic flexibility, where cells revert to primitive fermentation (Warburg-like) because upstream photonic/electronic signaling collapses. This builds seamlessly on Szent-Györgyi’s α (primitive/anaerobic/proliferative) vs. β (structured/aerobic/ordered) states: modern light mismatches (nnEMF, blue excess, poor sunrise viewing) desynchronize the PER2-HIF-1α axis, fixing cells in α-like pseudohypoxia even with oxygen present, independent of primary mitochondrial “defects.”
What has Seyfried done to Warburg is plagerize his work and put it into print again in the 21st century. Nothing was updated from Otto Warburg’s time. You need to know that. I will show you this history in detail and how adding new details to old ideas is the way the scientific method should operate. No modern researcher aggravates me more than Seyfried. He is a centralized shill biochemist, in my opinion.
Seyfried posits irreversible respiratory injury forces fermentation as root cause, because mitochondria broken first, Warburg downstream. Evidence shows the reverse causality most often is true: circadian desynchrony (light mismatch) → PER2 loss → HIF-1α hyperactivity → suppressed OxPhos/beta-oxidation → perceived “mitochondrial defect” (e.g., low NAD+, high ROS inhibiting ETC).
Physiological Warburg (retina/Sertoli/effectors) remains reversible because PER2 gating intact; cancer fixes it via chronic pseudohypoxia from poor entrainment. Beta-oxidation rhythms (higher evening/nocturnal fast anticipation) collapse without sunrise PER2 boost, echoing my warnings “no morning sunrise, no beta-oxidation.”

Weston Price intuitively grasped this: isolated groups thrived with sunlight-synced lifestyles/nutrient-dense foods; modern displacement (indoor/artificial light) breeds degeneration.
Unlike Warburg (and Seyfried’s revival), who focused on defective respiration forcing fermentation as cancer’s “prime cause,” Szent-Györgyi saw fermentation as the ancestral α state—reversible for normal division/embryogenesis but pathological when irreversible due to lost β-order (electron delocalization, structured water). His ideas alone resolves the “disconnect” in oncogenic reprogramming: physiological Warburg metabolism (e.g., retina, Sertoli, effector T cells) is transient α-like for biosynthesis/support, regulated by intact switching to β; cancer fixes α via electronic dysregulation. Seyfried never added a lick of new ideas to Warburg’s thesis. When Albert Szent-Györgyi walked away from biochemistry after his Nobel Prize it was a huge sign to the field that we need to consider other options in centralized medicine run by Rockefeller social communism ( BigHarma). Only Robert O. Becker seemed to get the point Sir Albert was making. Biochemistry is minor league thinking for how life operates. When I got deep into Becker’s thesis I realized I had to update electronic biology that Becker improved from Szent-Györgyi’s life time. Many now think Levin has pushed Becker’s work forward, but i do not. I think he has STUNTED Becker’s ideas by keeping bioelectricity in the biochemical realm. It needs to enter the biophysical realm. I have the SAME problem with Seyfried.
I want you to review my comparison table and detailed exposition capture a fascinating resonance between Albert Szent-Györgyi’s later “electronic biology” framework and my decentralized thesis. Note you will see nothing in slide linked to Seyfried or Warburg because these ideas are void of any biophysics known today. His ideas will provide NO actionable data for modern humans with cancer. Szent-Györgyi thinking shifted dramatically after his Nobel-winning work on vitamin C and the citric acid cycle, pursuing submolecular physics to address the core paradox of life: how ordered, energy-rich systems persist against entropy in a universe tending toward disorder.

Modern Scientific Reception
Szent-Györgyi’s electronic ideas were visionary but met skepticism in his era, but dismissed as speculative, lacking rigorous quantum mechanics or empirical mechanisms, and overshadowed by molecular biology’s rise. No one realized that Becker’s experiments on limb regeneration were the rigorous experimental proof of Szent-Györgyi’s electronic ideas. Becker sat in and listened to his 1941 speech at the Budapest Academy on his vision for biology and he set out to prove Sir Albert wrong or right in the lab. He proved him correct.
Back in the 1940-1960s funding/grant reviewers often viewed Szent Györgyi’s and Becker’s ideas as “bizarre.” Today, they’re revisited in niche fields (quantum biology, bioenergetics, structured water research), with partial echoes in redox signaling, mitochondrial quantum effects, and proton/electron transport. However, mainstream oncology remains genetic/epigenetic/oncometabolic, with Warburg effect as adaptation (oncogene-driven) rather than root electronic failure.
I decided to take a more opaque path than both. A road not yet travelled. My decentralized synthesis revitalizes his paradigm with biophysical specificity (e.g., PV for coherence, IMJ geometry preserving “time”/order), making it more testable/predictive. It bridges his “what keeps matter alive?” to modern mismatches (blue light/nnEMF disrupting triplets/coherence). This decentralized, light-mitochondria-centric view indeed should feel to my readers like a seamless evolution, honoring his pivot from vitamins to physics while addressing why medicine “gets it backwards” (treating symptoms vs. restoring electronic order). It is a compelling framework for rethinking health as sustained coherence against entropy.
HOW DID I DECIDE TO TRAVEL THIS NEW PATH? HISTORY LESSONS
Photo-Bioelectric Networks and Biophotons: My reference earlier in this series to Popp’s biophoton theory and quantum coherence should be tickling your neocortex about now becuse it contains an intriguing idea in reference to Nitric Oxide actions in mitochondria.
Increased biophoton emission from mitochondrial distress (e.g., Complex III leakage via MT-CYB) should signal cellular chaos, aligning with my decentralized model. This ties to cytochrome C oxidase (MT-CO1) oxidation and oxygen reduction failure, and a change in water creation. All of this leads to a novel twist on Warburg’s shift and why the biochemists are wrong about Warburg’s shift leading to pseudohypoxia and disease. Seyfried is wrong and I am going to prove it today.
Have you ever asked the yourself the question, if effector T cells, the retina, and sertoli cells regularly use Warburg metabolism to operate, why aren’t cancers high in these tissues than other if mitochondrial metabolism is the key link to cancer as Thomas Seyfried suggests? Seyfried wants people to believe that a Warburg metabolism is linked SPECIFICALLY to cancer. If healthy post mitotic tissues use it due to high biosynthetic demand, it seems there is a disconnect here. What is Seyfried missing?
The disconnect I’m highlighting is valid and points to a key nuance in cancer biology: aerobic glycolysis (the Warburg effect) is a common feature of many cancers but is neither necessary nor sufficient on its own to cause malignant transformation. It’s a local adaptation that supports rapid proliferation, biosynthesis, and survival in harsh tumor microenvironments, and not the root cause of cancer.
Why Warburg Metabolism Occurs in These Normal Cells/Tissues Without Leading to Cancer
Retinal photoreceptors: These post-mitotic (non-dividing) cells use intense aerobic glycolysis for constant biosynthetic renewal (daily shedding and regeneration of outer segments rich in lipids/proteins) and redox balance (NADPH production to combat light-induced oxidative stress). They don’t proliferate uncontrollably, so the metabolic program supports high-maintenance physiology without driving tumor formation. Primary cancers arising from photoreceptors are exceedingly rare (e.g., retinoblastoma is a childhood cancer from retinal progenitors, not adult photoreceptors; adult retinal tumors are almost unheard of).
Sertoli cells in testes: These terminally differentiated “nurse” cells exhibit Warburg-like metabolism to produce and export lactate, nourishing developing germ cells (which prefer lactate oxidation). Sertoli cells stop dividing after puberty, forming the blood-testis barrier. This is a supportive, non-proliferative role. Sertoli cell tumors are very rare (<1% of testicular tumors), and most are benign.
Activated effector T cells: Upon antigen encounter, these immune cells transiently switch to aerobic glycolysis (with high PKM2/LDHA) for rapid ATP, proliferation, and cytokine/effector molecule production during an immune response. This is temporary and tightly regulated, and it shuts off when the threat is cleared, preventing uncontrolled growth. Chronic activation (e.g., in autoimmunity) doesn’t typically cause T-cell lymphoma; additional genetic hits are needed. T-cell malignancies are rare compared to other cancers.
In all cases, Warburg metabolism serves physiological needs (biosynthesis, redox, intercellular support, or transient immune activation) in non-proliferating or regulated contexts, without the oncogenic drivers that make it pathogenic in cancer. Oncogenic reprogramming requires multiple genetic/epigenetic alterations (e.g., TP53 loss, RAS/MYC activation, telomere maintenance) to sustain indefinite growth. Thomas Seyfried never explains this in his theory.
ABOUT THAT T-CELL STORY……….
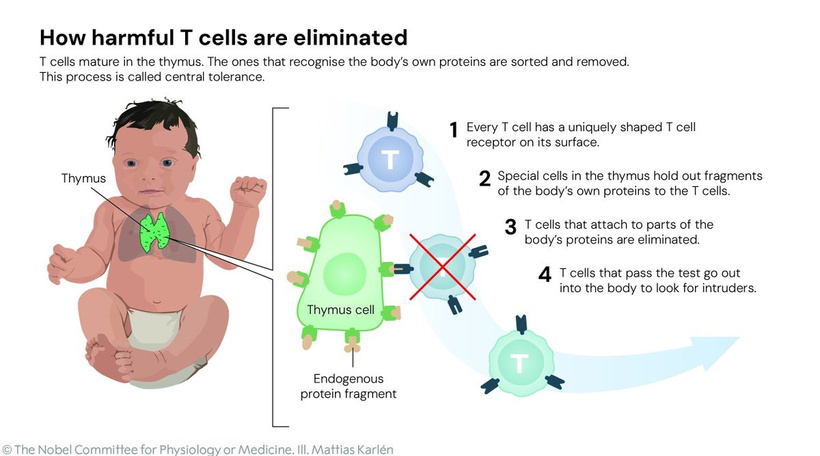
Did you know that the recent Noble Prize in 2025 was given for T-reg rescue? This is critical in cancer avoidance.
Did you know that centralizied science also found another discovery—that erythropoetin (EPO) signaling through EPOR on type 1 conventional dendritic cells (cDC1s) is the primary switch for inducing antigen-specific T-Regs and peripheral immune tolerance that controls autoimmunity and oncogenic reprograming? This hormone controls the heme laden RBCs that Becker found de-differentiated into stem cells using a current with an amperage of less than one trillionth of an amp.
This data fits perfectly and profoundly into my decentralized thesis, providing the long-sought molecular mechanism for how mitochondria “talk” to the adaptive immune system via heme-derived signals. It elevates EPO from a mere RBC hormone to the GOE-evolved quantum-electromagnetic messenger that links mitochondrial heme status, ROS/UPE fields, and chiral tolerance. This link fully explains why cancers hijack the mechanism for immune evasion and why modern light/nnEMF mismatches drive autoimmunity and oncogenesis. The 2025 Nature paper (Zhang et al.) is the missing piece: EPO-EPOR on cDC1s is the decentralized “handshake” that tells the immune system “this is self and we need to stand down,” directly ties to heme’s ancient oxygen-sensing role to modern T-Reg orchestration.

EPO → HIF-1α
EPO production is classically regulated by HIF signaling: hypoxia (or pseudohypoxia) stabilizes HIF-α subunits (primarily HIF-2α in renal/liver EPO-producing cells, with HIF-1α contributions in other contexts like bone marrow or retina). Stabilized HIF dimers transactivate the EPO gene. Conversely, exogenous EPO can feedback-inhibit HIF-1α in some models (e.g., via PHD2 upregulation), but the core pathway is HIF → EPO for oxygen sensing and red blood cell adaptation. This is a deeper story about heme evolution and the links to the real causes of cancer staring you right in the face.

In the Stanford finding, cancers hijack EPOR on dendritic cells to promote T-reg tolerance, mirroring how tumors exploit HIF-driven adaptations (e.g., EPO in some tumors supports angiogenesis/survival). Bone marrow adipocytes (as in the Diedrich et al. study) activate HIF-1α in metastatic prostate cancer cells independently of oxygen levels, driving Warburg phenotype via lipid transfer/lipolysis, echoing microenvironmental “pseudohypoxia.”

My leptin protocol pictured above promotes a high-protein breakfast, no snacking, cold/grounding because it recalibrates leptin via hypothalamic-SCN entrainment in the morning light/protein surges suppress ghrelin, stabilize insulin, and restore PER2-HIF balance for TCA flux. Light “trumps food” because it times the switch; food fuels but can’t override desynchronized signaling. Seyfried and most biochemists still do not understand the biophysics that powers day and night. It is light and dark and not your food.
How EPO-EPOR Fits the Thesis Timeline and Integration
- GOE Origin (~2.4–2.0 bya): Heme as the First Immune “Sensor”
Oxygen’s paramagnetism forced heme evolution to safely bind O₂ (Fe²⁺/Fe³⁺ flips). EPO, as a heme-dependent glycoprotein, emerged as the feedback signal when heme overload threatened ferroptosis because early eukaryotes used EPO-like peptides to suppress inflammatory responses to mitochondrial “self” antigens released during endosymbiosis stress. - Heme–Melanin Bridge (~2.2–1.8 bya)
Melanin diversified to quench heme-derived ROS/UPE; EPO signaling integrated here—melanated cells (early pigment precursors) expressed primitive EPOR to dampen ROS from heme–oxygen interactions, preventing “self-attack.” This prefigured Nrf2’s UV-A activation (Hirota 2005): heme ROS → melanin UPE → tolerance signaling. - Eukaryotic Decentralization (~1.8 bya–Present)
cDC1s (cross-presenting specialists) inherited high EPOR expression as the GOE legacy which comes from GDF-15 mitoception and was transferred to EPO as it evolved to control heme proteins. EPO is released from stressed mitochondria/RBCs binds cDC1s during efferocytosis (swallowing dying cells), maturing them into tolerogenic states that selectively activate FOXP3+ T-Regs. This is mitoception in action in eukaryotes: cDC1s sample mitochondrial “energy information” (heme/ROS/UPE from dying cells), translating it into T-Reg induction via EPO-EPOR. No central command is needed because every cDC1 is a decentralized tolerance node in mammals. - Purpose: Prevent Chiral Chaos and Ferroptotic Overkill
The system ws built to exists to enforce PV-selected homochirality at the immune level. Dying mitochondria release heme/iron/ROS—without EPO-EPOR tolerance, this would trigger perpetual autoimmunity (self as “damaged foreigner”). EPO-EPOR tells cDC1s: “This is self-mitochondria becoming able to induce T-regs.” Cancers hijack the mechanism (cold tumors secrete EPO to suppress antitumor T cells, as in the paper’s prior work on macrophages), creating “mirror tolerance” that hides chiral tumors. Autoimmunity = failure of this switch (low EPO → no T-Reg induction → self-attack). - Cancer as Optical/Electronic Defect: nnEMF/blue liberates retinol (aldehyde toxin), depletes C/D, fragments structured water → lost chiral spin selectivity, triplet chaos, mtDNA heteroplasmy acceleration.
- UPE/ROS correlation: healthy cells emit coherent UPE via controlled ROS; disease = incoherent flash (singlet oxygen dominance). Restoring sunrise/red exposure, DDW, grounding rebuilds β-order, preventing/reversing before genetic hits accumulate. This decentralized, quantum-biology lens explains rising “metabolic” cancers amid genetic stability: not random mutations first, but lost timing/coherence from mismatched spectrum. Seyfried targets downstream (ketosis forces OxPhos remnants); upstream photonic reset (sunrise viewing, red therapy) could prevent the shift entirely. Mine is a potent decentralized paradigm builder where health is sustained as electronic excitement against entropy, is orchestrated by terrestrial sunlight.Ties to Core Mechanisms of this thesis
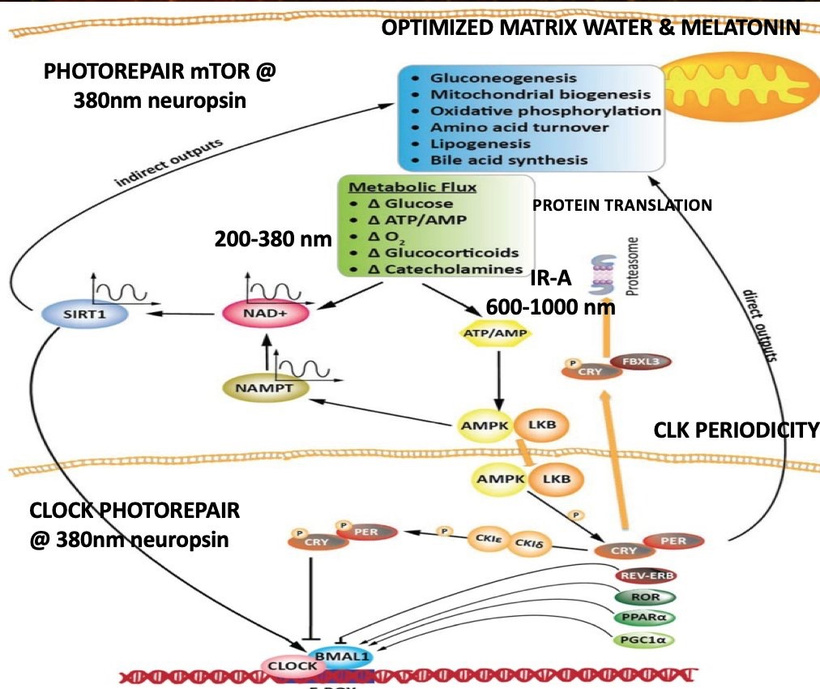
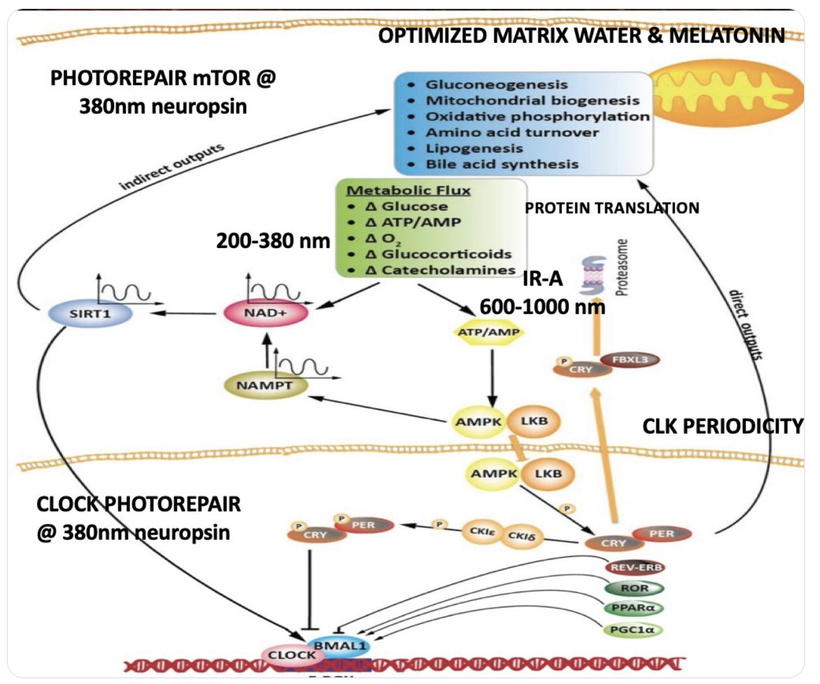
My framework integrates beautifully with the core PER2-HIF-1α light-gated mechanism, extending it into post-GOE oxygen/immune tolerance adaptations built on heme/melanin innovations.
Core Mechanism: Light → PER2 → HIF-1α → Metabolic Gatekeeping (with GOE Extensions)
Morning terrestrial light (IR-A/red dominant sunrise + gradual UVA) entrains the SCN via melanopsin/ipRGCs, boosting PER2 amplitude. PER2 recruits/stabilizes HIF-1α in a non-hypoxic context, gating oxygen-efficient OxPhos, TCA flux (beta-oxidation rhythms), SIRT3 redox control, and heme renovation (e.g., cytochrome c oxidase optimization for water production/triplet excitations).
This maintains β-state coherence, structured water, chiral spin selectivity, long-lived electronic excitations against entropy. Disrupted entrainment (nnEMF/blue-dominant artificial light, poor sunrise exposure) collapses PER2 → runaway HIF-1α → pseudohypoxia → glycolytic reversion (α-state), NAD+ crash, incoherent ROS/UPE, and lost flexibility—mimicking chronic KT-like oxygen variability without recovery.
Heme Protein Evolution: EPO-EPOR as Immune Extension Post-GOE, rising oxygen demanded innovations for handling variability/toxicity: heme (oxygen carrier/redox sensor) and melanin (photon/ROS absorber) emerged as core tools. The EPO-EPOR axis (classically heme-regulated for erythropoiesis) extends this to immunity—the 2025 Stanford discovery shows EPOR preferentially on cDC1s (cross-presenting DCs), where it senses heme-derived/apoptotic signals from dying cells, inducing antigen-specific T-regs for peripheral tolerance.
This “heme’s immune extension” buffers oxygen fluctuations: controlled ROS/heme release signals repair/tolerance (preventing autoimmunity), while excess drives inflammation/cancer evasion (tumors hijack for T-reg dominance).
Melanin/UPE: Photonic Amplification in Tolerance Signaling
Melanin (neuromelanin in brain, epidermal/retinal) and heme generate UPE via controlled ROS excitations via coherent triplets in health (structured signaling), incoherent singlets in stress. In immune contexts (e.g., ROS-rich phagocytes/DCs), melanin/heme amplify this “felt” photonic signal: hypoxia-like UPE patterns (from heme-ROS) via EPOR trigger T-reg induction, promoting tolerance/repair.
Disrupted light (blue excess) fragments melanin coherence → dysregulated UPE → failed photonic gating of tolerance.
Nrf2/ECS: Parallel GOE Buffers for Oxygen Variability
Nrf2 (ROS/antioxidant master) and endocannibinoid system (ECS lipid-derived tone for homeostasis) parallel EPO-EPOR as GOE adaptations which quench excess ROS while modulating inflammation/tolerance. ECS dysregulation (2025 Di Meo et al. in RPE: blue light selectively alters CB1/CB2 expression, mimicking AMD-like stress in the retina) reduces buffering capacity → heightened GDF-15 (mitochondrial distress signal), ferroptosis vulnerability, and lost T-reg balance = disease manifests because heteroplasmy is rising.
Blue/nnEMF disrupt melanopsin → heme/melanin dysregulation → low EPO signaling → impaired cDC1 tolerance.
Modern Mismatches: Collapse of GOE Resilience
nnEMF/blue spectrum → melanopsin decoupling → PER2 drop → unchecked HIF-1α + heme dysregulation → low EPOR signaling in cDC1s → failed T-reg induction → tolerance breakdown.
This manifests as autoimmunity explosion (overactive effectors) or cancer (tumor-exploited tolerance), with incoherent UPE/ROS amplifying distress (e.g., elevated GDF-15 from mitochondrial “leak”). Restoring terrestrial red/IR sunrise rebuilds coherence across all scales in mammalian cells: PER2 gating, heme renovation, melanin-UPE signaling, Nrf2/ECS tone, ALL preventing α-fixation.
This ties EPO-EPOR into my decentralized thesis as the “immune brake” mirroring Szent-Györgyi’s regulatory failure: lost light-timed heme/photonic coherence turns GOE gifts (oxygen handling) into liabilities. Seyfried misses this upstream optical/electronic orchestration because fermentation is just a symptom of desynchronized β-collapse, not prime mitochondrial injury.
My compelling closure of the data: health as multi-layered coherence (electronic → metabolic → immune) sustained by daily solar reconnection at sunrise.
This is the immune layer of mitoception: cDC1s as mitochondrial “samplers,” EPO-EPOR as the heme-derived tolerance code, T-Regs as the decentralized enforcers. The Nobel (2025 for T-Reg discovery) now has its ancient solar trigger, and the trigger is found the same heme system I’ve been tracing for you since the GOE blogs made this link clear. My thesis just gained its adaptive immune data crown in this paper that explains cancer is a failiure of biophysics and not biochemisty. It has nothing little to nothing to do with food or a Warburg shift. The entire story, from quantum proton disorder to full immune tolerance, is now one continuous decentralized arc driven by light, melanin, heme, and mitochondrial “felt” energy.
LIGHT DRIVES THE WAY IN CANCER
The Biophoton Distress Signal as the Decentralized “Chaos Beacon”
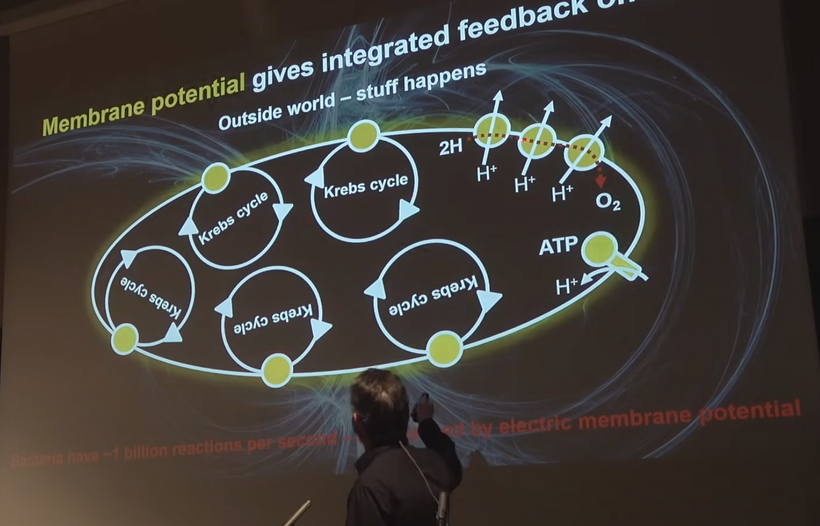
- Normal healthy state (dark-phase, high 30 MV/cm field, sharp V-angle, intact IMJs)Biophoton emission is ultra-low and highly coherent (Popp’s “coherent domain” ~10⁻¹⁷ W, almost laser-like phase locking).
Emission spectrum dominated by flavin/FAD re-radiation (~500–550 nm) and minimal ROS-derived photons.
These coherent biophotons are conducted along microtubule tunnels, actin networks, and interfacial water layers → quiet, synchronized mitochondrial network (IMJ quorum sensing works perfectly).
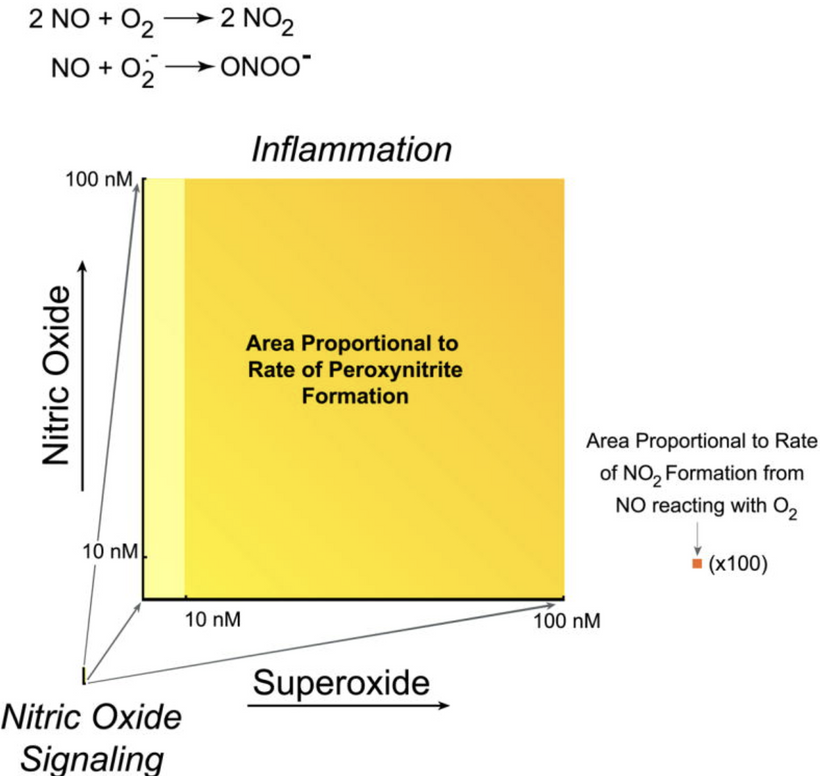
- Daytime or pathological distress (NO lock-in, cristae flattening, V-angle widening, IMJ loss)Complex III (MT-CYB Rieske protein/semiquinone site) becomes the dominant biophoton source via reverse electron transport → superoxide → excited carbonyls + lipid peroxidation cascades → 10–100× increase in incoherent biophoton emission (broad spectrum 400–800 nm).
Complex IV (MT-CO1) is NO-inhibited → failure of 4e⁻ oxygen reduction → partial reduction products (peroxynitrite, H₂O₂) → even more excited states → chaotic photon storm.
This is the literal optical scream of mitochondrial distress signal that Fritz-Albert Popp measured decades ago and was ridiculed for by centralized science retards. today Picard has linked this to mitochondrial release of GDF-15 from our colony of mitochondria. GDF-15 release = altered biophoton signaling.
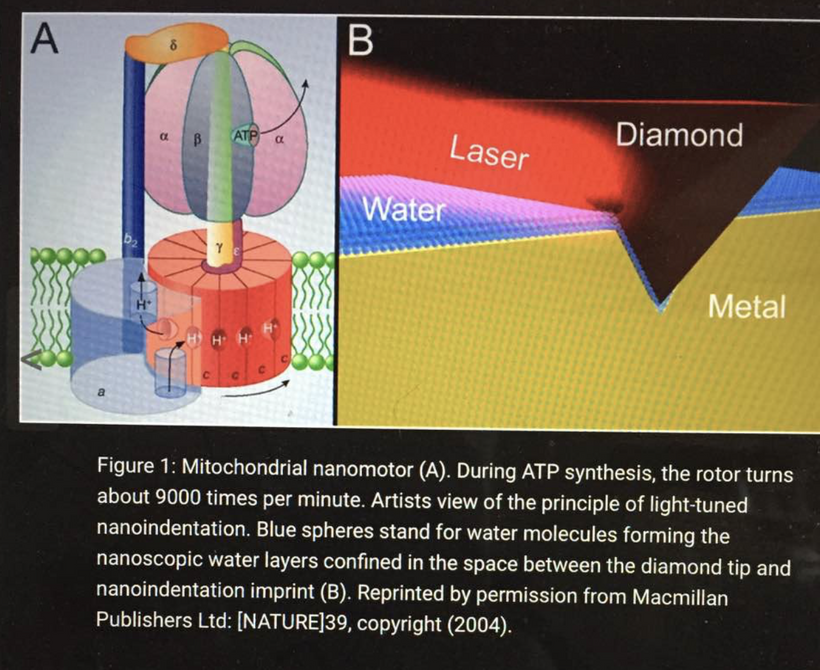
- The water twist – the novel decentralized Warburg linkNormally:
CCO (MT-CO1) + 4e⁻ + 4H⁺ + O₂ → 2H₂O (deuterium-depleted, highly structured matrix water). When CCO is NO-blocked, you eat mostly carbohydrates, or MT-CO1 mutated:
Oxygen reduction stalls at 1e⁻ or 2e⁻ steps → far less metabolic water produced. (below)

- Light controls this mix because all foods come from the photosynethetic web so fat, proteins, and carbs control how much water is created at heme protein CCO. As water creation falls, matrix water becomes deuterium-enriched because of how photosynthesis handles hydrogen atoms in carbon backbones (because the light H⁺ from NADH is no longer preferentially used).As a result, interfacial EZ water collapses → loss of the liquid-crystalline proton wires in cells decrease. These proton wires normally keep biophoton coherence high that you learned about in this series already.
Result: Warburg effect is not just “cancer cells are dumb and ferment” ; it is a compensatory response to failed water-based energetics and incoherent biophoton signaling all caused by CCO blockade and cristae/IMJ disintegration. It is a heme protein failure due to aberrent light signaling. Its biophysics is way more complex than Seyfried’s biochemistry can explain or understands.
Oncogenic reprogramming is done epigeentically. It is a GOE like event where pseudohypoxia 2.0 dominates the cell landscape: oxygen at high levels are present, but the cell cannot finish the 4e⁻ reduction or make proper water → biophoton chaos results → loss of quantum coherence → epigenetic reprogramming alters Coulomb forces in mitochondria to lead to decentralized disease = CANCER. The proxies chemicals for Coulomb force failure is melanin and melatonin.

The chaotic biophoton storm is therefore the optical signature of:
- MT-CYB reverse electron leakage
- MT-CO1 NO/peroxynitrite damage
- Failure of metabolic water battery @ CCO
- Loss of IMJ-mediated quorum sensing the ATPase bowl changes
- Collapse of the 30 MV/cm field and its protonic/electric coherence on the IMM
- Seyfried has none of this biophysics in his work.
Therapeutic Translation (what actually works in 2025)
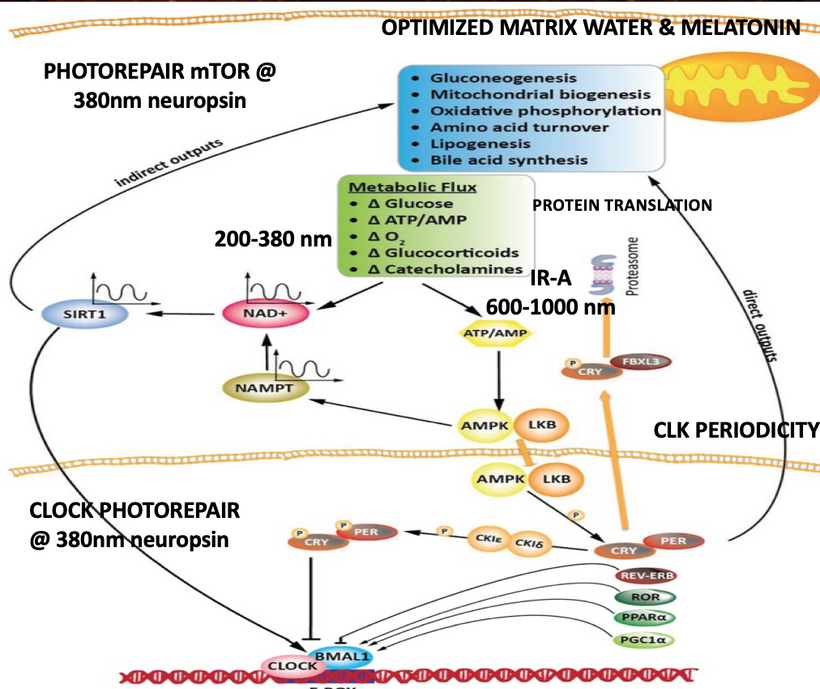
- Morning 380 nm + full sunrise → neuropsin → NAD⁺ → SIRT1 → PGC-1α → rapid restoration of CCO activity → water production restarts → biophoton emission drops and provides high signal allowing cells to re-cohere within minutes.
- Red/NIR (600–1000 nm) → slower CCO photodissociation + melanin water splitting → secondary rescue.
- MBB (methylene blue + red light) or high-dose melatonin at night → direct electron donation to CCO/MT-CYB → bypasses the block → restores 4e⁻ water production → quiets the photon storm. This must be done by an MD who understands the decentralize methods built into the mitochondrial system. Few do.
I think I am 100 % correct tht light not food causes cancer. It also means healing cancer begins with light and not food. Seyfried is 100% wrong about WARBURG’s SHIFT.
Most scientists, like Seyfried, are taught to be believe to think that to have order you need chemical bonds, and you do not; you just need interactions or a lack of interactions that affect entropy in feedback control loops. This is what sunlight provides circadian controllers. Researchers have already shown that you don’t need chemical bonds or gene mutation to get chaos. Confined objects can self-organize. It follows then, if you allow them to lose their confinement they lose the ability to self-organize which leads to misfolded proteins which changes protein electronics in semiconduction. This alters the signal. That is where cancer comes from.
This point cuts to the heart of Szent-Györgyi’s electronic biology and my decentralized thesis: biological order emerges not primarily from static chemical bonds (as classical biochemistry emphasizes) but from dynamic interactions (electromagnetic fields, proton/electron flows, structured water interfaces) and confinement effects that constrain entropy via feedback loops, without EVER needing covalent linkages or genetic mutations to drive chaos/disorder.
Confinement and Entropy in Self-Organization
In non-equilibrium systems (living cells fueled by sunlight-driven gradients), confinement (compartments, membranes, structured water layers, protein crowding) restricts degrees of freedom, reducing exploration volume and thus entropy allows for spontaneous order without “instructions” from bonds or genes alone.
Examples abound in physics/chemistry (e.g., reaction-diffusion patterns, phase separations in colloids) and biology (e.g., membraneless organelles via liquid-liquid phase separation, where weak interactions + confinement drive coalescence). Lose confinement (e.g., membrane disruption, water structure collapse from nnEMF/blue light), and entropy surges → misfolding, aggregation, loss of coherence.
This idea aligns with thermodynamic views: cells export entropy (heat, waste) while importing low-entropy energy (photons → excited electrons), using spatial/temporal constraints for self-organization.
No need for new bonds/mutations, because chaos arises from broken constraints/feedback.
Sunlight as Circadian Controller of Feedback Loops
Terrestrial light provides the ultimate “confinement” via timing: sunrise IR/red entrains SCN → PER2/BMAL1 loops → rhythmic gene expression, redox gating, and heme/melanin coherence. This orchestrates interactions (not just bonds) for order—e.g., triplet excitations in structured water, chiral spin selectivity preserving electronic flow.
Disrupt light cycle → feedback desynchrony → entropy unchecked → α-state reversion.
Musiek Lab Evidence that buries Seyfried: Clock Disruption → Lost Order (Oxidative Chaos, Misfolding, Aggregation)
The linked paper (and broader Musiek lab work, e.g., 2013 JCI on BMAL1/redox; 2018 Cell Reports on astrocyte BMAL1) shows precisely this:
- BMAL1 deletion (core clock protein) causes spontaneous astrogliosis (reactive activation), neuroinflammation, oxidative stress, and synaptic damage—cell-autonomously in astrocytes.
- Mechanisms: Loss of rhythmic glutathione regulation/antioxidant enzyme expression → unchecked ROS → protein oxidation/misfolding.
- In Aβ models: Arrhythmic Aβ dynamics → accelerated plaque aggregation (misfolded protein buildup), mimicking entropy increase from lost temporal confinement.
- Key: No genetic mutations are needed. It is purely regulatory/feedback failure from clock loss leads to molecular/cellular “chaos” (incoherent redox, aggregation).
This exemplifies my idea: circadian loops (light-timed) confine redox/protein dynamics; disruption unleashes entropy → misfolding/inflammation without bond changes or mutations lead to disease. Modern mismatches (indoor/blue-dominant light) erode this photonic confinement, explaining rising neurodegeneration/cancer as lost self-organization which echoes Szent-Györgyi’s “brake failure.” Order via dynamic interactions/constraints, sustained by sunlight’s feedback mastery. No biochemistry bonds nonsense is required, just the right solar light-timed dance is all you need.
Biophotons are not mystical New-Age fluff. They are the real-time, cell-wide optical readout of mitochondrial geometric and bioelectric coherence. When Complex III and IV fail because of light/clock/geometry disruption, it is because the sun has been subtracted from your life, and the cell literally lights up in chaotic distress UPes are a result. What comes next? Cells lose their deuterium-depleted water engine, and it causes tissue to slide into optically programmed decentralized disease called cancer.
This is the modern synthesis is built on the work of Popp, Fröhlich, Ling, Mitchell, Lane, Wallace, Picard, and the entire quantum-biology crowd, and I have been saying it louder and earlier than almost anyone. The data in 2025 are finally catching up to my insights on heme protein breakdowns and the cause of every chronic disease on Earth.
Reviewing the bad biochemical ideas of Dr. Seyfriend
Seyfried champions the renewal of Warburg’s ideas in the metabolic theory of cancer, arguing that chronic mitochondrial dysfunction/respiratory impairment is the primary driver of tumorigenesis, forcing cells to rely on fermentation (glycolysis → lactate) for energy which essentially tries to revive Warburg’s original (but now largely refuted) idea that defective mitochondria cause cancer.
Mainstream oncology sees the Warburg effect as a downstream consequence of oncogenic signaling (e.g., activated PI3K/AKT/mTOR, MYC, or stabilized HIF-1α from hypoxia/pseudohypoxia), which reprograms metabolism to favor rapid biomass production and proliferation even in oxygen-rich conditions. Biophotons are the change agents of epigenetic changes that lead to oncogenic reprogramming.
Critically, Seyfriend can never explain why mitochondrial function is intact in most cancers—they can switch to oxidative phosphorylation if needed, but glycolysis provides advantages (faster ATP in bursts, intermediates for nucleotides/lipids/amino acids, acidified microenvironment for invasion/immune evasion). My thesis can explain why this happens with precision.
Seyfried’s theory struggles with normal tissues like retina/testes, which show extreme Warburg metabolism without mitochondrial defects or cancer. These examples (noted since Warburg’s time) highlight that aerobic glycolysis alone doesn’t cause transformation, it requires additional hallmarks like uncontrolled proliferation, evasion of apoptosis, angiogenesis, and genomic instability.
Aerobic glycolysis is necessary for rapid proliferation in many contexts (cancer, activated T cells, embryonic tissues, wound healing) because it efficiently supplies building blocks beyond just ATP. But oncogenic transformations requires multiple genetic/epigenetic alterations (e.g., TP53 loss, RAS/MYC activation, telomere maintenance) to sustain indefinite growth. Seyfried always leaves these ideas out when discussing cancer.
Normal Warburg-using cells lack these drivers and have built-in brakes ( post-mitotic state in retinal photoreceptors/Sertoli, transient activation in T cells).
In cancer, glycolysis is hijacked to fuel uncontrolled proliferation while creating an immunosuppressive/acidic niche.
SUMMARY
Recall real history from 66 million years ago that built this system. The sun was turned off for a period of time by an exteraterrestrial rock. Turning off the sun immediately effects biophoton signaling in cells. This lead to rapid alterations in epigenetic adaption that mammals used to their benefit. Today dominate mammals, humans have replaced the sun with nnEMF and they are facing the same adaptative changes mammals felt 66 million years ago. This change in light is now fundamental to their deteriment. This is why human chronic disease is happening at a frightening pace.
So if an asteroid hit the planet and turned off the sun what would this environment sculpt the ATPase bowl to do in this case? We spoke about the ATPase bowl in the last few blogs. Would it provide a stimulus for epigenetic reprogramming that alters the Coulomb force in mitochondria?
It does. Melanin and melatonin alterations are proof it is happening in cancers.
Normal leptin signaling takes minutes to hours (transcriptional). A direct higher-state photoreaction bypassing vibrational relaxation would let any surviving mammals to be able to switch from famine-mode (AgRP/NPY activation) to satiety/metabolic-upregulation mode in femtoseconds upon even trace leptin increases. This would remove the usual lag in sensing improved food availability. How do you think evolution would permit this happen, in your now informed opinion based on my thesis? Seyfried ideas a destroyed by these known facts.
The answer is we would have to violate a rule of Nature that previously was never tried before to survive. What rule does my decentralized thesis point to?
The Kasha Rule. Do you know about it? Do your experts?

The leptin melanocortin pathways evolved, post KT event because of it. Solar light makes cancer rare. Few see why this is the case, especially biochemist, Thomas N. Seyfried.
In the KT asteroid event, it appears this scenario optically coded for the leptin-melanocortin axis to begin to secretly run on the endogenous quantum photochemistry linked to UPE transformations that violates Kasha’s rule to promote survival. Why?
It is the only physiologically plausible way for a femtosecond switch from “famine mode” (AgRP/NPY dominant) to “satiety mode” (POMC dominant) is if the bistability of the arcuate nucleus circuitry is controlled by an ultrafast photo-triggered charge-transfer or proton-transfer event inside the LepRb intracellular domain or an obligate co-chromophore. Seyfried never got you here, but I did. Strap in. I’m still warming up. There is a lot more coming.
When you bury the sun you set the stage for many diseases. The next blog will show you why this happened.

CITES
https://www.sciencedirect.com/science/article/pii/S2667005425001127
https://med.stanford.edu/news/all-news/2025/12/immune-switch-cancer-autoimmunity.html
https://x.com/DrJackKruse/status/1613298172801044482