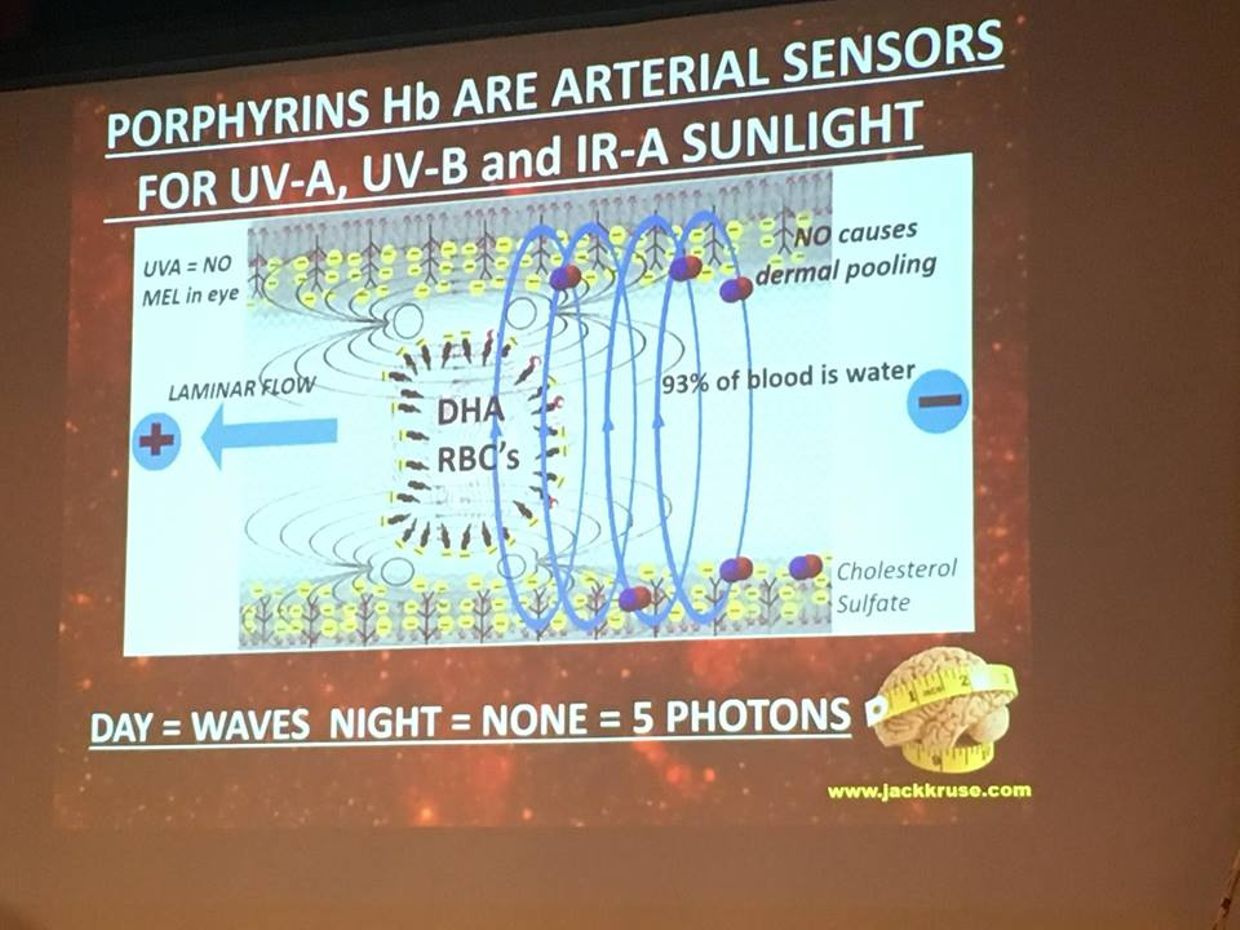
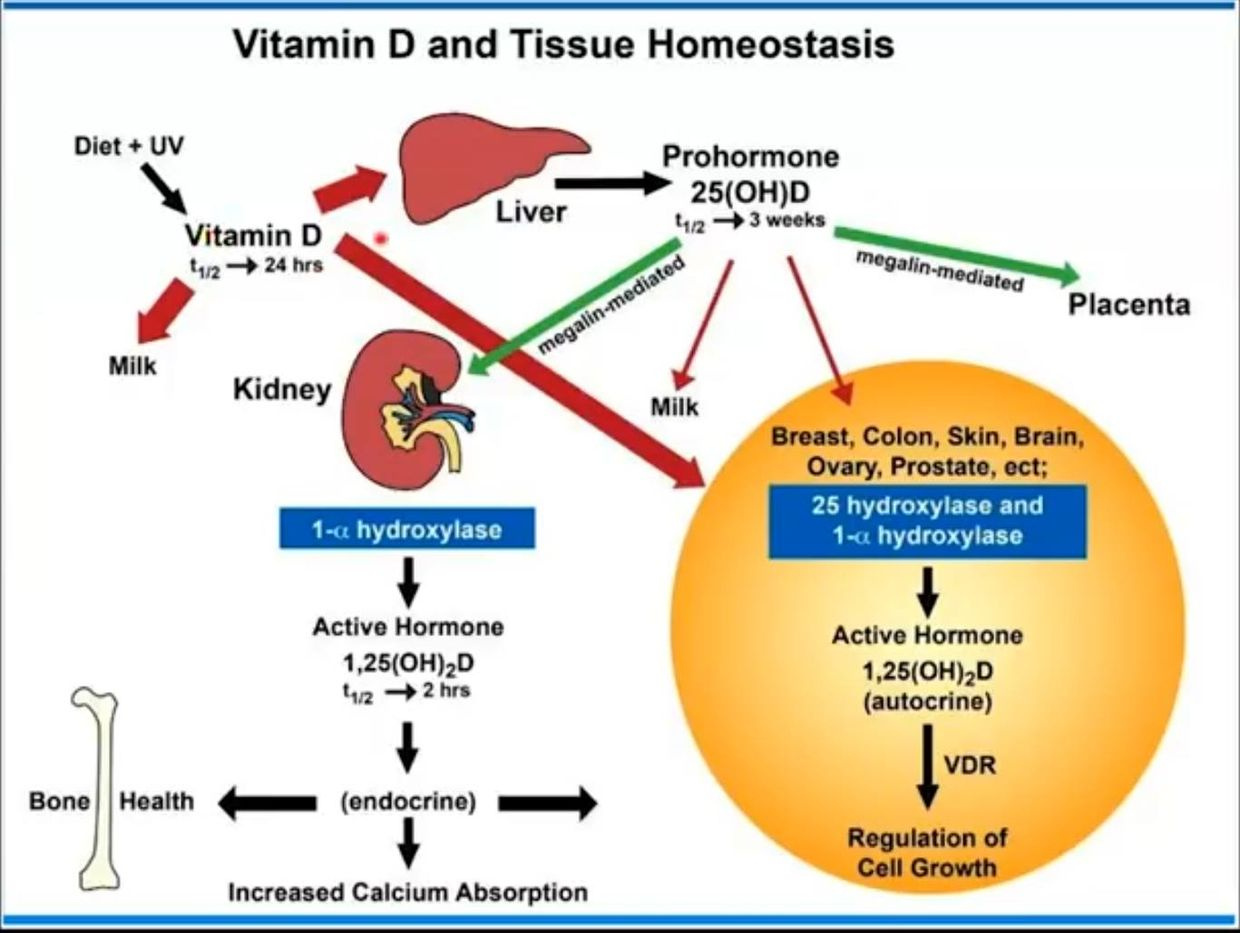
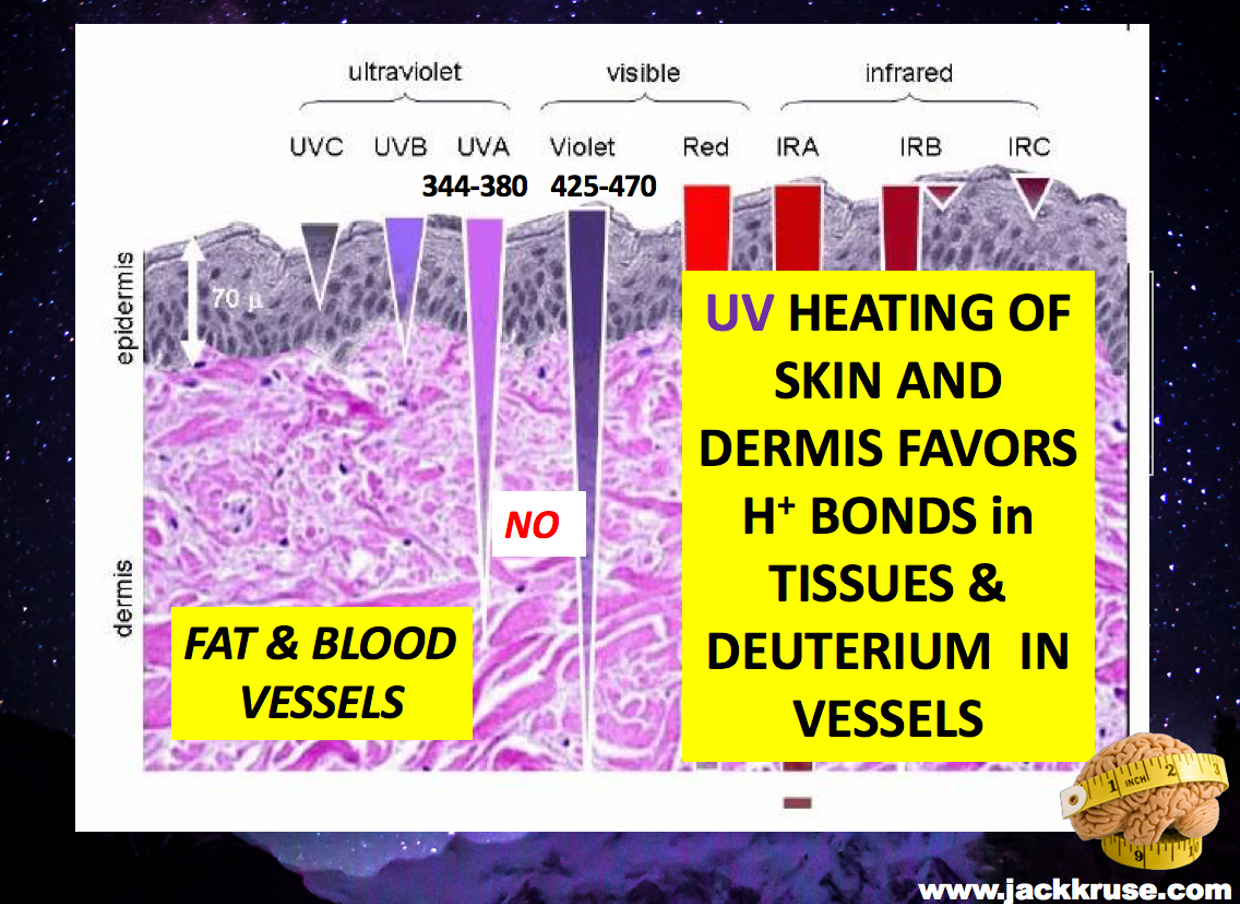
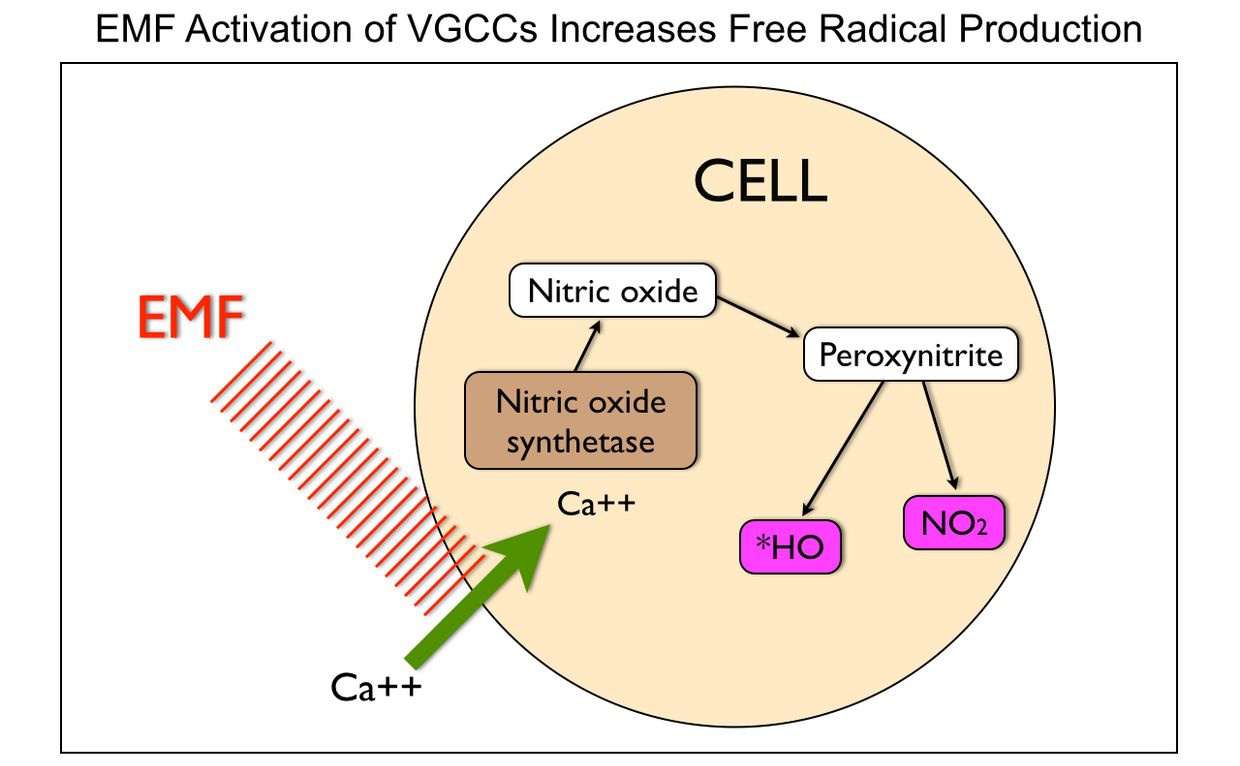
Few of us realize the liquid crystals in us create time using light. Time and space are modes by which we think and experience reality but neither are conditions in which we live on Earth. We live by swinging on the spiral arm of a galaxy constantly moving through space .
Ever since the derivation of the theory of relativity, physicists have thought they have done reasonably well in understanding time; however, biologists are still waiting for a comprehensive theory of timing in living systems: they remain waiting for a corpus of “laws” describing how cells and organisms can precisely initiate and terminate processes at specified times because they are focused on the nuclear genome and not the mitochondrial genome. Time perception is critical to mitochondrial biology but it is not especially important to the nuclear genome because the energy provided by the mitochondrial genome allows time to manifest in living things. This deficiency in biology is particularly acute in developmental biology, where complex mechanisms of various paces and durations must be orchestrated to solve huge developmental problems such as the one faced by the fertilized egg: how to become an organism using energy/information and electric and magnetic field to direct and power morphogenesis.
Animal development is, in fact, nothing but time. From the cell cycle to the beating of the heart, our own lives are composed of a multitude of microscopic and molecular oscillations. For developmental biology, the study of causal relationships implies the examination of two time points: inducing the cause and looking at the effect. In QED, cause and effect is blurred because of the uncertainity principle. In biology, cause and effect “appear” to be linked because of how time is perceived. We are generally ignorant of the temporal rules governing this transformation. Light and aromatic amino acids form those foundations. Whereas the rules of a Swiss watch involve a timing mechanism that uses a uniform unvarying tempo to translate rapid oscillations (seconds) into longer periods of time (hours), the rules of the developmental clock of animals are much more complex because they use light and light is non linear in the violet part of the visible spectrum. The animal clock is made up of a variety of counting mechanisms that follow varied temporal rules at different frequencies and often run in parallel without any “apparent” interaction with each other, because their linkages are via molecular resonance. The goal of the developmental clock is not simply to mark off time, but to integrate and unify the myriad temporal signals received from throughout the organism.
However, the frequencies and amplitudes of most, if not all, developmental clocks are robustly determined within the same species, suggesting that these are genetically encoded devices being linked and controlled by resonance instead of being produced by the system itself, as an “emergent feature.” I have always believed time is an emergent property of life because time emerges from the complexity of light the force carrier of the electromagnetic force. Today, the “old paradigm of biologic time (Franz Hallberg) is dominant in the published literature, but as Max Panck said, things change in science one funeral at a time. Morever, if the “belief” is true that time is encoded in our DNA (it’s not), what are the genetic balance wheel, hairspring, and gears, and what are the specialized tools that are needed to elucidate the intricate workings of the genome? There are many possibilities to consider here, but I will discuss two.

The first one concerns clocks generated by mechanisms in “trans”, generally based on the postulate made in the early 1950s by Alan Turing (mathematician, philosopher, and inventor of the concept of the computer), who argued that the diffusion of substances with activation, repression, and retroactive potentials can generate intrinsic oscillations in space and time. The literature has begun to decipher some of these ideas and more are likely to come in the near future.
The second possibility involves mechanisms in “cis”, using the linearity of our genetic material itself, our chromosomes, to measure time; an admittedly elegant solution, but one that severely lacks both experimental and theoretical support. Chronobiology is stuck on these two options because they do not realize that time is a superposition of many possibilities. This is why they ignore the mitochondrial genome. This genome creates many possibilities when small changes occur to it. When we disrupt it with anesthesia timing is completely altered, we observe it and we measure it, but we do not understand the implications for man or his diseases. This will change with the coming age of quantum biology. The old guard expected some help from the human genome sequencing and related high-throughput technologies, but they got stonewalled because it destroyed constructs that we inherited from Watson and Crick. I reject most of their thesis about biology.

Modern epigenetics data has blown up much of Darwin’s thesis from 1859 but few biologist will admit it today because it is akin to removing the binding from a book. All organizing principle are afloat now in molecular genetics because of the new insights of epigenetics. So is science and life. Every end is a new beginning in understanding and learning with respect to nature’s wisdom. Darwin’s supporters today are still trying to hold the theory together but the concept of timing and energy is 100% tied to energy collection and generation and distribution in the cell. They have no Earthly idea that energy and information processing are coupled by light.
There is little doubt that the mechanisms in ‘trans’ will be advantageously studied in this context. Time-lapse proteomics and metabolomics, adapted to minute amounts of material such as cohorts of less than a thousand cells, would certainly show significant recurrences and, perhaps, allow the temporal algorithms underlying developmental clocks to be uncovered. But trends in protein or metabolic movement can just describe the cellular processes and it won’t explain it. The explaination will take a mind filled with innovated thoughts to link them back to the fabric of nature’s laws. Today’s developmental chronobiologists believe that a reductionist approach to clock mechanisms will give us the key answers. In fact, we just gave three people a Nobel Prize recently for discovering the biologic molecules involved in the circadian mechanism. For example, the tightly regulated timing in the successive appearance of whiskers on the nose of a rodent would likely be deciphered if we could microdissect the whisker field like a chess grid, and submit each square to full protein content analysis.
On the other hand, the challenge in identifying ‘cis’ processes would be to use several species showing graded variations in clock parameters, such as in the frequencies of the same clock, and to find correlations with the “cis organization” of DNA segments through a large-scale approach. Such correlations, if any, could reflect the existence of linear time-measuring devices, from the mere duration of transcription to more complex processes such as progressive transitions in the chromatin state. However, that is easier to say than to do; a genuine “casse-tête” for bioinformaticians.

I believe based upon what we know today about epigenetics, it is seriously pretentious to try to extract the fourth dimension, time, out of our DNA/RNA structure when we still lack complete understanding of the other three dimensions of our world. But the human quantum computer does not need complete data sets to jump to conclusions. That is what innovation and imagination are useful for in complex problem solving. This science is the privilege and the difficulty of working with embryos: The spatial construction can be understood only in the light of time. The irony of this idea, is that light is what creates time.

This concept continues to trip up biologists because they are ignorant of the queerness of the physics of light. Similarly, evolutionary genomics will likely teach us what happened during phylogenetic times, and our ontogenic clocks may be revealed by global comparative analyses.

Chronomics is on the way, but only the unripe mind will think it is the be all end all in the discussion of what time really is.
Time allows us to see what we have become, even when we miss it. The liquid crystals inside of us build this library. Time is erased for some of us, because we cannot use sunlight and water to build the crystals properly to manage our time. Sometimes we erase people and family, and walk away from the shadows creeping up on us. Sometimes you must give up on people, not because you do not care, because they do not care enough for themself by making the correct choices.