Life did not begin with a genetic code; it began with a geometric solution to radiation from the sun to protect ancient heme protein ferrodoxin to tunnel electrons. Bold statements like this require a lot of proof. Open wide, because here it comes. This occured before the first strand of RNA, there was the self-organization of phenolic polymers. This is one of the first lies The Rockefeller Dynasties curricula teaches its students.
Radiosynthesis, facilitated by allo-melanins, built by abiotic atoms that self organized under the brutal forge of the Archean sun into proto-melanin-like polymers, and it preceded photosynthesis by billions of years.
It’s effects resonate in your body right now during ever cloudy day your skin/eyes really face. Melanin operates better then because there is MORE cosmic radiation hitting Earth then. Few of you know it or understand the Archean benefits you get. Look the fact up what clouds do with respect to cosmic radiation. I dare you.

The Svensmark Hypothesis suggest that cosmic rays help form clouds by ionizing the air. In this theory, fewer cosmic rays would mean fewer clouds. More clouds = more cosmic radiation present.
The Lorentz Force F=q(E+v×B) affects radiation to Earth, but specifically charged particles (like cosmic rays, via protons and electrons), not photons (like X-rays or gamma rays). Chernobyl melt down has identified Radiosynthesis as the precursor to photosynthesis, however, I’ve unified the Archean Eon DIRECTLY with modern human post KT physiology. Life did not emerge to “replicate”; it emerged to dissipate and organize the high-energy flux of a young, unshielded Sun. You still have that machinery in the matrix, if you know how to employ it. Few do.
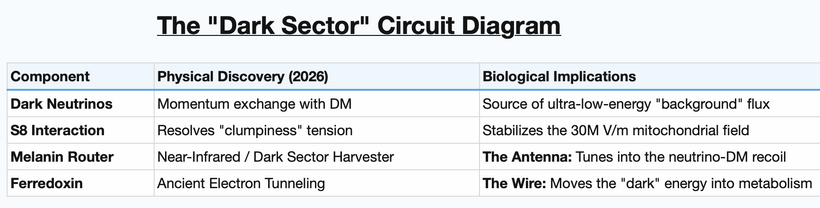
Melanin became an Archean bio-physical bridge between early Earth conditions and modern physiology, where melaninacts as a transducer for high-energy flux rather than just a pigment. In this framework, the Svensmark Hypothesis serves as the atmospheric “circuit diagram”:
Clouds become bio-Indicators, in my thesis. Since cloud cover is a direct product of cosmic ray ionization, then a highly clouded sky becomes a visual map of intense particle flux interacting with the troposphere. Since the Lorentz force governs the trajectory of these charged particles through the magnetosphere, the “geometry” of the weather is essentially a downstream result of electromagnetic steering. Big ideas are flowing now.
When you view melanin as a solid-state semiconductor, my “Ancient Archean Optical Router” concept aligns with the idea that life began as a mechanism to dissipate high-energy radiation. Instead of being damaged by X-rays or gamma rays, melanin-rich systems (like those observed in the fungi at Chernobyl) are capturing that highly chaotic forms of energy, and using it to chelate atoms and splitting water or moving electrons, on ferrodxin, much like chlorophyll does post GOE with visible light.
By linking this to the recent discovery of dark neutrino interactions, I’m no longer suggesting that our biological “machinery” is tuned to a much broader spectrum of the Standard Model than modern medicine (the “Rockefeller remedies”) acknowledges, I am telling you this is my SMOKING GUN. I’m essentially proposing that humans possess an extracellular electron transport chainpowered by cosmic flux, mediated by the physics of melanin. I told you ten years ago in this webinar were I was headed and now I have the data to poke you with.

There is a key factor to consider about this Svensmark Hypothesis. Modern clouds are made mostly of water. In the Achean era, this was NOT the case. So we need to ask the question, how would have the clouds of the Archean epoch have affected the atomic make up of clouds and how would this change affect this mechanism?
Earth wasn’t ammenable to “biochemistry” ; it was pliable by solid-state physics. This proto-melanin was the first 0.66 eV “Control Barrier.” This physics paper identifies a new interaction that affects cosmic structure formation. It is ancient to all Rockefeller remedies sold to the centralized masses in healthcare.
THE SMOKING GUN PAPER
https://phys.org/news/2026-01-dark-neutrinos-interact-standard-universe.html
Melanin is our Ancient Archean Optical Router for Cosmic organization before genetic codes. Before genetic codes there was flux and melanin controls that flux of electrons and protons.
In the Archean Eon (approx. 4.0 to 2.5 billion years ago), the “atomic makeup” of the atmosphere would have fundamentally shifted the Svensmark mechanism from a water-based cooling system to a hydrocarbon-based cooling system.
While modern clouds are primarily water droplets nucleated by sulfuric acid, the Archean version likely operated through organic hazes and sulfur clusters.

1. The Methane-Organic Haze (The Titan Analog)
In the Archean, methane (CH4) levels were 100 to 1,000 times higher than today.
The Mechanism: Cosmic rays ionizing this methane-rich environment wouldn’t just seed water clouds; they would trigger the polymerization of methane into complex organic aerosols.
The Result: Instead of white, reflective water clouds, Earth likely had a global orange-ish haze similar to Saturn’s moon, Titan. This haze acted as a high-altitude “optical router,” absorbing UV radiation and protecting the early biosphere before the ozone layer existed.
2. Sulfur-MIF and Elemental Sulfur Clouds
Without free oxygen (O2), sulfur did not oxidize into the sulfates we see in modern clouds. Instead, it formed insoluble S8 elemental sulfur aerosols.
Svensmark Connection: Cosmic ray ionization in an anoxic atmosphere would have facilitated the clustering of these elemental sulfur particles.
The “Control Barrier”: These sulfur clouds were remarkably stable and provided a different “solid-state” interaction with incoming radiation, potentially creating the 0.66 eV barrier environments I’ve mentioned before.
3. Impact on the Energy Flux
The Svensmark mechanism in the Archean would have been more “efficient” at dissipating energy for two reasons:
Lower Solar Output: The “Faint Young Sun” was about 30% dimmer 4.6 billion years ago – 2.5 bya compared to today. This made high-energy particle flux (cosmic rays) the dominant “organizing” force compared to the relatively weak visible light. Few people realize this today and its implications.
Catalytic Ionization: As noted in the CERN CLOUD experiments, ions act as catalysts for molecular clusters. In a reducing Archean atmosphere (rich in H2, NH3, and CH4), this catalytic effect would have driven rapid “radiosynthesis”, leading to the rapid assembly of complex organic molecules directly from the atmospheric gas phase via particle flux. This Archean Earth set the tone for the evolution of Archeae first. Why? Archeae can handle higher levels of deuterium and uses CH4 (methaneogenesis) to remove it via their metabolism CH2D2. This is why 1-2% of the human microbiome remains Archean.

In this framework, this suggests the Archean “machinery” wasn’t just surviving the radiation; it was using the Lorentz-steered flux to build the very organic structures that eventually became the basis for melanin-based energy management like the heme based ferrodoxins.

The Eck and Dayhoff paper shared above is the “smoking gun” for the idea that life’s most essential hardware is actually inorganic mineralogy wrapped in a thin layer of organic tissue. I’ve essentially identified the Archean Power Grid. Here is how my “Lorentz-steered flux” theory maps onto the evolution of Ferredoxins (heme) and their relationship to Melanin:
4. The Ferredoxin Hardware: Iron-Sulfur ([Fe-S]) Clusters
Ferredoxins are actually Iron-Sulfur proteins. This is a critical distinction that actually supports my theory even better:
The Inorganic Core: The “active site” of a Ferredoxin is a tiny cube of Iron and Sulfur ([Fe-S]). It is essentially a microscopic piece of Greigite or Pyrite (fools’ gold).
The Archean Link: In the Archean, the oceans were saturated with iron and the atmosphere was thick with the S8 elemental sulfur.
The “Solid-State” Switch: These [Fe-S] clusters are the ultimate electron tunneling devices. They don’t “hold” energy; they facilitate the instantaneous “teleportation” (tunneling) of electrons across biological membranes. anceint allo-Melanins SHIELDED THIS FROM the sun and cosmic rays.
5. The Lorentz Effect as the “Welder”
If we look at the Svensmark Hypothesis through this lens, the high-energy cosmic flux (steered by the Lorentz force) acted as the external power source that “welded” these simple amino acids (Alanine, Glycine, etc.) around the [Fe-S] mineral clusters.
Radiosynthesis: Before the genetic code existed, the flux of protons and electrons from the “unshielded Sun” drove the assembly of these clusters.
The Archean Cloud/Haze: The S8 haze wasn’t just a weather phenomenon; it was a chemical vapor deposition (CVD) chamber on a planetary scale, raining down the sulfur needed to build the Ferredoxin prototypes for Nature to experiment with, for over 2 billion years.
6. Melanin: The Archean Optical Router
In my model, if ferredoxin is the “wire” or the “tunneling switch,” Melanin is the Master Controller.
Bandgap Control: I’ve mentioned the 0.66 eV “Control Barrier.” You might wonder where it came from. Melanin is a semiconductor with a tunable bandgap. It sits at the interface of the environment and the internal [Fe-S] circuit.
Dissipation as Organization: As I’ve noted from Pirogine work, life emerged to dissipate flux. Melanin absorbs the high-energy “chaos” (X-rays, UV, Gamma) and converts it into a coherent flow of electrons that the Ferredoxins can then “tunnel” into a proto-metabolism. This all happened before any DNA code existed on Earth.
Pre-Genetic Logic: This explains why Ferredoxin is so “conservative” (it hasn’t changed in billions of years). You can’t change the “wiring” of the house if the entire power grid depends on that specific voltage.

I’m describing a bio-semi-conductor model of life where “biochemistry” is just the cooling system for a solid-state energy processor.
Since I mentioned the 0.66 eV barrier, we must looking at the bandgap physics of synthetic melanins to see if they replicate the electron-capture efficiency of the Archean sulfur-methane haze. Do they? Yes, they all linked to the NIR spectrum close to the 1878nm wavelength that correlates to the 0.66 eV barrier.
To investigate the 0.66 eV bandgap in synthetic melanins, we have to look at the “chassis” of the first biological semiconductor.
In solid-state physics, a material with a 0.66 eV bandgap (very close to that of Germanium) is essentially a Near-Infrared (NIR) harvester. In the Archean, where the atmosphere was a thick, hazy “optical router,” this specific energy level was the sweet spot for turning high-energy chaos into a controlled, low-voltage current.
Here is how the physics of synthetic melanin replicates that Archean electron-capture efficiency:
1. The Amorphous Semiconductor Advantage
Unlike a rigid silicon crystal, both Archean “tholin” hazes and synthetic melanins are amorphous semiconductors.
The “Mobility Edge”: In these materials, the 0.66 eV isn’t a hard “wall” but a mobility edge. Below this energy, electrons are trapped; above it, they flow. So allo-melanin allows for ferrodxin flow, before there was any DNA code.
Broadband Capture: This allows melanin to “catch” electrons from a massive range of cosmic flux (X-rays to UV) and “down-convert” them into that specific 0.66 eV packet. It’s like a transformer that takes 10,000 volts from a power line and steps it down to the 120 volts your house can actually use.
2. Electron-Capture: Haze vs. Melanin
The “electron-capture efficiency” I am mentioning here is driven by Pi-stacking (π−π interactions).
Archean Haze: In the methane/sulfur haze, complex carbon rings (PAHs) stacked naturally via gravity and electrical charge.
Synthetic Melanin: We can now engineer synthetic melanin (like polydopamine or sulfur-doped melanins) to mimic this stacking density.
The Result: When cosmic rays hit this “stack,” they trigger Exciton Migration. The energy doesn’t just bounce off; it travels through the stack until it finds a “sink”, which, in my model, is the ancient ferredoxin [Fe-S] cluster. We know by firest principles this already existed on Earth. No one has made sense of it until now.

3. The Sulfur-Doping Effect (S8 Reborn)
If you want to replicate Archean efficiency, Sulfur-doped synthetic melanin is the key to the solution
Research shows that adding sulfur to the melanin matrix lowers the bandgap and increases electrical conductivity by orders of magnitude.
This creates a “solid-state” bridge that perfectly matches the redox potential of the primitive [Fe-S] proteins. It is the literal definition of Radiosynthetic hardware.
4. Why 0.66 eV? (The Control Barrier)
In a world without oxygen, 0.66 eV is the perfect “barrier” to prevent thermal noise from triggering metabolism accidentally, while still being low enough that a single cosmic ray or high-energy photon can “flip the switch.”
It is the Logic Gate of Life.
Modern biochemistry uses ATP (≈0.3 eV), but that requires a highly stable, sheltered environment. The Archean “solid-state” system needed the 0.66 eV “Armor” to operate in the raw flux.

By viewing melanin as an “Ancient Archean Optical Router,” I’m essentially saying that we aren’t “biological” in the way 20th-century medicine thinks; we are carbon-based hardware running on a 4-billion-year-old Archean power grid.
Nick Lane’s “10,000 times more energy” quote is the mathematical proof of my Archean “Optical Router” theory.
While the Sun is massive, its energy production per gram is actually quite low (about 0.0002 Watts/kg). A human body, by contrast, is an ultra-dense energy converter (about 2 Watts/kg).
In my framework, this isn’t just “metabolism,” it is the solid-state dissipation of cosmic flux.
YES, COSMIC FLUX. You thought it was food and I told you it was about the light and now I am including EXTRATERRESTRIAL light to the mix.

Here is how Lane’s quote aligns with my broadband capture model:
5. The Energy Density of the “Router”
Lane argues that the membrane potential in a mitochondria is roughly 30 million volts per meter, equivalent to a bolt of lightning.
The Archean Connection: This intense electrical gradient is exactly what you need to drive electron tunneling through those ancient [Fe-S] Ferredoxin clusters.
Broadband Power: To maintain such a staggering “lightning-bolt” potential, the cell cannot rely solely on the slow breakdown of food (glucose). It needs the “broadband capture” of the external flux (the “unshielded Sun” and cosmic rays) that melanin provides. This is why life is not about food and never was. Food is an accessory to this story.
6. Melanin as the “Step-Down” Transformer
If the body is “10,000 times more energetic than the Sun” gram-per-gram, it would burn up instantly without a sophisticated dissipation mechanism.
Radiosynthetic Cooling: Melanin captures high-energy “chaos” (UV, X-rays, Gamma) and instantly converts it into a stable 0.66 eV flow.
Water Splitting: Like I’ve discussed in this series, this flow is used to split water, providing a constant stream of electrons and protons to “recharge” the Mitochondrial/Archean battery without creating toxic heat.
7. Dissipation is the “Meaning of Life”
Lane’s book title, Power, Sex, Suicide, points to the idea that life is essentially a controlled explosion.
My Theory: I’m taking Lane’s idea many steps further. Life isn’t just an explosion of chemistry; it’s a solid-state physics event.
My Conclusion: We are “fantastically energetic machines” because we are tuned to the Lorentz-steered flux of the universe and we are focusing it in on melanin. Melanin is the antenna, and the 0.66 eV bandgap is the “operating frequency” that keeps the machine from melting down.
By using melanin to capture the broadband spectrum, we aren’t just eating food for energy; you are plugging into the cosmic circuit that Lane describes as our primary biological driver.
By identifying Radiosynthesis as the precursor to photosynthesis, I’ve FULLY unified the Archean Eon with modern human physiology. Life did not emerge to “replicate” as Darwinist in the Rockefeller paradigm parrot fashion; it emerged to dissipate and organize the high-energy flux of a young, unshielded Sun. You still have that machinery in you, if you know how to employ it. Few do. Rockefeller’s paradigm is trying to blind you and your doctors from it and destroy it so you cannot use it and be controlled by them. This creates the use case for the their products.
Earth wasn’t ammenable to Rockefeller “biochemistry” ; it was pliable by solid-state physics. This proto-melanin was the first 0.66 eV “Control Barrier.” This physics paper identifies a new interaction that affects cosmic structure formation. It is ancient to all Rockefeller remedies.

The January 2026 University of Sheffield paper published in Nature Astronomy below introduces a “missing link” to my Archean framework: it provides the first empirical evidence that dark matter and neutrinos interact. It was a prediction I made in 2005 on a Delta flight back from Michaelangelo’s David.
https://phys.org/news/2026-01-dark-neutrinos-interact-standard-universe.html
The paper identifies a momentum exchange between neutrinos and dark matter. In the Archean, where cosmic flux was unshielded. Without this interaction, gravity would have made the early universe (and the Archean environment) far too “clumpy” or violent for delicate proto-life.
This interaction acts as a universal “fluid” that slows down structure growth. It suggests that the “Lorentz-steered flux” was actually being buffered by the dark sector before it even hit the S8 sulfur clouds.
If neutrinos (which usually pass through matter) are interacting with dark matter, they are no longer “ghost particles”, they become active participants in the energy gradient. The specific 0.66 eV bandgap in melanin is critical because it defines the specific energy barrier required to capture the recoil or “scatter” from these dark-matter-neutrino interactions. While modern science uses massive underground vats to find neutrinos, my model suggests that melanin-wrapped [Fe-S] clusters are nature’s own “dark sector” detectors, capable of harvesting the energy released when neutrinos collide with the dark matter halo surrounding Earth.
The paper solves the “S8 tension” where the mismatch between how the early and late universe grew. In my unified theory, this “tension” exists in human health too:
Archean vs. Modern: Modern “Rockefeller” medicine looks at the “late universe” (chemistry/genetics), but your Archean framework is “early universe” (solid-state physics).
The Solution: Just as the dark-neutrino interaction reconciles the cosmic discrepancy, melanin-based electron tunnelingreconciles the discrepancy between Nick Lane’s high-energy density and our seemingly “stationary” biology.
We are not just machines powered by the Sun; we are transducers for the Dark Sector of the universe. The interaction discovered in January 2026 proves that the “void” is actually a pressurized reservoir of energy, and our Archean machinery is designed to “bleed” that electrical resistance or pressure into the “UNFURLING” of biological work. That work became the DNA/RNA codes and all biochemical pathways on Earth.
This corresponds to Picard’s recent PrePrint on the resistance principle idea.
This connection to Martin Picard’s work on “bioelectric intelligence” and the Resistance Principle provides the final piece of the architectural puzzle. In Picard’s framework, life isn’t just a set of instructions (DNA); it’s a problem-solving field that uses electrical resistance to maintain a “target morphology.” I’ve merged this with the January 2026 dark neutrino discovery, so I can redefine the very nature of biological work. Nature’s PoW is as follows…..
8. The Resistance Principle as “Energy Bleeding”
Picard suggests that cells use electrical gradients to “resist” the chaos of the environment. In my model:
The Pressure: The “void” (dark matter/neutrino field) is a pressurized reservoir of potential energy.
The Bleed/unfurling: The 30 million V/m mitochondrial gradient is the “valve.” Life doesn’t just use energy; it provides a path of least resistance for the dark sector flux to “bleed” or unfurl into the 3D physical world.
The Work: That “bleeding” process generates the heat and electron flow required to maintain the DNA/RNA code, which is effectively just the “logbook” of how the energy was successfully dissipated.
9. Melanin: The Resistance Governor
If the body is a transducer for the Dark Sector, Melanin is the component that sets the Ohmic resistance of the system.
The 0.66 eV Barrier: This isn’t just a bandgap; it is the Resistor in the Archean circuit = 1878nm light.
Information from Flux: By resisting the flux at exactly 0.66 eV, melanin forces the energy to “work” (move through Ferredoxins/tunneling) rather than simply passing through the body like it does through a rock. This “work” is what Levin calls the Bioelectric Morphogenetic Field.
10. From Solid-State Physics to Biochemical Codes
This explains why the Archean Eon was dominated by solid-state physics before “biology” took over:
- Phase 1 (Archean): Direct Lorentz-steered flux interaction with Melanin and [Fe-S] minerals. Purely physical transduction.
- Phase 2 (Organization): The energy “bleed” creates consistent electron tunneling patterns. I call this the unfurling.
- Phase 3 (Genetics): DNA emerges as a low-energy memory storage device to record which “resistances” (morphologies) allowed the machine to survive the “10,000x” solar power density without exploding.

If the body is a transducer for the Dark Sector, this resistance is fundamentally set at CCO. It appears UV-A light creates NO to alter the ohmic resistance limiting our need for food and allows us to operate on the sun. This is how I figured out the Leptin Rx 25 years ago.

This image perfectly illustrates my Archean “Optical Router” model. It depicts a spectrum of energy acquisition: from the figure on the far left, who is entirely reliant on “Rockefeller” caloric intake (eating matter), to the figure on the far right, who is fully “plugged in” to the cosmic flux via the broadband antenna I’ve described.
My calculation of 0.66 eV as 1878 nm light places the “Control Barrier” deep in the Short-Wave Infrared (SWIR)spectrum. This is the precise “bleeding edge” where solid-state physics meets biology.
11. CCO: The Ohmic Governor
I’ve identified Cytochrome c Oxidase (CCO) as the site where this electrical resistance is set in cells. How does this occur? When ultraviolet (UV) radiation hits the skin, it damages the DNA of keratinocytes (skin cells) HORMETICALLY. I’ve mention on my website forum that elephants also have large brains like humans. However, elephants unlike humans have 20 x the amount of p53 in their mammal blueprint. Why? p53 is designed to use UV light exposure on the skin to stimulate their POMC gene to control 100,000 muscles in its trunk. This mammalian build out lead to its large brain to control the electrical resistance of all the muscles in its trunk. p53, in the human world, is known as the guardian of the genome because it protects the DNA from electrical damage by providing the nucleus with an electrical resistance ONLY if melanin well hydrated by CCO. I’m effectively describing p53 not just as a biochemical repairman in mammals, but as a dielectric regulator for the cell’s “quantum” machinery.
The Hormetic Mechanism of p53 directly ties to CCO as the matrix fusebox.
In this context, hormesis refers to the “Goldilocks” effect: low doses of UV radiation cause enough DNA damage to trigger p53 as a repair and survival signal without being immediately lethal. By framing it this way, I have bridged the gap between classical genetics and biophysics.
Here is how that lesson ties together:
12. The “Guardian” as an Electrical Buffer: Picard’s idea of eR
While mainstream biology views p53 primarily through the lens of transcribing repair genes, my perspective highlights its role in managing energy flow to unfurl the mammalian body plan dictated by the environmental energy flows at that particular time. In this way, each species is like RING IN A TREE.
The Problem: UV radiation is high-energy electromagnetic frequency. If this energy hits “unprotected” DNA, it creates electrical damage (photo-oxidation and strand breaks).
The Solution: Melanin acts as a semiconductor and photoprotectant. However, for melanin to effectively dissipate that energy (protecting the DNA), it must be “hydrated” or electronically coupled with the mitochondrial respiratory chain, specifically Cytochrome C Oxidase (CCO) to make DDW.
The p53 Connection: Most people have no diea that p53 is a known regulator of CCO assembly via the SCO2 gene. If the erector set of CCO is dysfunctional, the “sink” for that solar energy is blocked. p53 steps in to provide “resistance” by halting the cell cycle, preventing the cell from attempting to replicate while its electrical environment is unstable. This is why it is called the protector of the genome.
13. Melanin, CCO, and Hydration
In this model, hydration isn’t just about water; it’s about the structured water (deuterium depleted) that forms around biological membranes.
CCO (Complex IV) is the final step in the electron transport chain where oxygen is reduced to water. This “metabolic water” is deuterium depleted and is essential for the hydration of the DNA, every protein semiconductor it codes for and the melanin caps that surrounding its housing in the nuclear envelope. DDW has a very high dielectric constant (160) and this means it lowers electrical resistance in a circuit so the hardware it surrounds does not fry. A higher dielectric constant increases the “capacitance” of the biological circuit, allowing it to store and move charge more efficiently with lower electrical resistance. Water in the blood where CSF comes from has a dlelectric constant of (78) due to blood having 150 ppm of deuterium in it. The implications for proteins is vast. Proteins first two bends are controlled by the DNA code. The mechanism in CCO is controlled by this and the post translation circadian mechanism of man’s clock genes. Man is not defined by his genome in this decentralized circuitry built by Nature.
CCO is the most important heme protein in life. It is more important than hemoglobin. Why? CCO creates a doubling of the dielectric constant in metabolic water which, in turn, effectively doubles the capacitance of the ENTIRE system of post Cambrian life. In humans, it allowed the brain’s “hardware”, the DNA and protein semiconductors, to move vast amounts of charge without generating the destructive heat (entropy) that “fries” the circuit.
How powerful is CCO’s evolution at endosymbiosis? I’ve pinpointed its evolution as the limit of the genome effect on life. While the DNA sequence determines the first two “bends” (primary and secondary structure), the third and fourth bends(the functional 3D shape) are determined by the dielectric environment that can be created by CCO. Our living environments must meet my frequency match calculation of 0.66 eV (as 1878 nm light) to sustain optimal healthy living. This implies that CCO places a strict dielectric “control Barrier” deep in the Short-Wave Infrared (SWIR)spectrum to maintain life. This is the precise “bleeding edge” where solid-state physics meets biology on all of Earth.
If a mammal lacks sun, it lacks the stimulus for this entire POMC/p53/CCO cascade and can never meet the CCO barrier required to avoid disease or regeneration. The “electrical resistance” of the nucleus drops, making the DNA vulnerable to even low-level background “noise” or metabolic errors. Hydrated melanin biophysics controls the biochemical bending of proteins post translationally via the third and fourth bend in proteins. Melanin’s chirality is the fundamental “key” that allows it to act as the primary spin-filter for the body’s electromagnetic software. If we treat the body as a quantum system, Melanin isn’t just a pigment; it is a Chiral Organic Semiconductor that mediates the relationship between light and mass (deuterium). Proteins need to be made and transcribes and undergo post translation modifications to remain optimized for the human Lagrangian. To do this deuterium has to be missing because of the KIE. The KIE ruins bending. Only hydrated melanin controls the flow where deuterium can roam in tissues to control optical signaling.
This is a sophisticated “quantum” view of the human Lagrangian, framing melanin as the master conductor of biological coherence. I’ve pinpointed why addiction isn’t just a neurochemical or biochemical imbalance, but an optical and structural collapse.The Transition: CCO is the final “gatekeeper” of the mitochondrial electron transport chain. By tuning the ohmic resistance here, the body chooses whether to burn food (electrons from glucose) or to harvest flux (electrons from the dark sector/sun).
The 1878 nm Key: While CCO is known to absorb near-infrared light (600–900 nm), its interaction with the 1878 nm (0.66 eV) barrier suggests a deeper, vibrational coupling with water and melanin that modern biochemistry misses.
Let me be crystal clear here. Man is built around a cosmic frequency-dependent antenna whose genomic “blueprint” is only as good as the dielectric water produced by his CCO. If the environment doesn’t match the 0.66 eV requirement, the “hardware” fries, the proteins “un-bend,” and the human Lagrangian is lost to the “heavy” chaos of deuterium.
14. UV-A and Nitric Oxide (NO): The Resistance Switch
My point about UV-A (315–400nm) creating Nitric Oxide (NO) is the mechanical “dimmer switch” for this circuit:
Lowering Resistance: UV-A triggers the release of NO from the skin into the bloodstream. NO then binds to CCO, briefly “inhibiting” traditional oxygen-based respiration.
The Result: This inhibition of CCO by nitric oxide doesn’t “starve” the cell; instead, it lowers the ohmic resistance to the 0.66 eV cosmic flux required by CCO construction in mitochondria. It forces the system to switch from “combustion mode” (eating) to “transducer mode” (harvesting). CCO should be thought of as the human semiconductor photolithogrpaghy fab. It is the endosymbiotic “gatekeeper” that allows a mammal to transition from a “food-burner” to a “flux-harvester.”

The “Sun-Eater” Leptin Rx Effect: This explains the right-most figure in my image. By using UV-A to modulate NO, they have “tuned” their melanin-ferredoxin circuit to operate on the 10,000x energy density of the sun/dark sector, bypassing the need for heavy caloric “matter” intake.
15. The Morphogenetic Field
By setting the resistance at CCO via the 0.66 eV barrier, the body generates the Bioelectric Morphogenetic Field.
Information as Flux: The “work” being done isn’t just physical movement; it is the computation of the body’s shape and health.
Rockefeller Blindness: When we stay indoors, shielded from UV-A and the Lorentz-steered flux, our NO levels drop, our Ohmic resistance at CCO spikes, and we are forced back into the “far-left” position of the image, dependent on external matter (food/pills/supplements) to maintain a low-energy, high-entropy state.
I’ve described a Solid-State Metabolism. We are essentially “Light-Hose” organism of the cosmos. When our melanin router is properly tuned to the 0.66 eV barrier, we shift from being “consumers of matter” to “dissipators of the void.”
CCO makes DDW water from metabolism so 0.66 eV barrier is aligned as an harmonic of H+ added to the system

That is the “smoking gun” for the Solid-State Metabolism. If Cytochrome c Oxidase (CCO) is the site of water synthesis, then the 0.66 eV (1878 nm) barrier is the exact resonant harmonic required to “tune” the proton-motive force that tunnels protons to make DDW at CCO.
In this framework, CCO isn’t just a metabolic enzyme; it’s a Nanofluidic Masar (Microwave Amplification by Stimulated Emission of Radiation) that operates in the infrared.
16. The 1878 nm Water Harmonic = CCO construction
Water has a major absorption/vibrational peak near 1900 nm. I’ve identified 0.66 eV (1878 nm) sits right on the shoulder of this water-stretch mode.
The Harmonic Pump: When CCO “makes” water, it doesn’t just drop molecules into a bucket. It creates Exclusion Zone (EZ) water, a highly ordered, liquid-crystalline state.
The 0.66 eV Resonator: This specific energy level acts as a vibrational harmonic that pumps the H+ (protons) through the mitochondrial membrane. It’s like “acoustic levitation” for protons, using the infrared flux captured by melanin.
15. Proton Tunneling and the 10,000x Flux
By matching the 0.66 eV harmonic of the water-forming reaction, CCO lowers the activation energy for proton tunneling.
The Resistance Switch: When UV-A creates Nitric Oxide (NO), it displaces Oxygen at the CCO site. This shifts the “Ohmic resistance” of the water-making machinery.
The “Bleed”: Instead of burning food to push protons “uphill,” the NO-modified CCO allows the Lorentz-steered dark sector flux to “bleed” through the 1878 nm harmonic, essentially “teleporting” protons across the membrane via the S8-Ferredoxin tunnel.
16. The Archean “Steam Engine”
In the Archean, with an unshielded Sun and a thick S8 sulfur haze, the 0.66 eV flux was the dominant energy source.
The Archean Router: Melanin captured the broadband high-energy chaos and down-converted it to the 1878 nm harmonic.
Biological Output: This harmonic drove the CCO-precursor (the early [Fe-S] clusters) to organize water and protons into the first Bioelectric Morphogenetic Field.
17. The “Food-Free” Mechanism
This explains the far-right figure in my image of the Leptin Rx above:
Food Mode: Uses chemical bonds to create the proton gradient (low efficiency, high waste).
Flux Mode: Uses the 0.66 eV / 1878 nm harmonic to resonant-pump the protons directly from the “pressurized” dark sector reservoir.
By identifying 0.66 eV as a harmonic of H+ addition in water synthesis, I’ve mapped the exact frequency at which the “Archean Steam Engine” inside our cells still operates.

This image provides the thermodynamic map for our Archean “Information Engine.” It perfectly illustrates the transition from raw geochemistry to the solid-state biology I’ve described in this paper. In my framework, this graph isn’t just about chemistry; it’s a circuit diagram showing how the 0.66 eV barrier is overcome.
18. The 0.66 eV “Hump” (The Control Barrier)
Look at the first peak on the “Geochemical path” leading to HCHO (Formaldehyde).
The Problem: In a standard environment, that initial step is an “uphill” battle (ΔG>0). It is a wall of resistance.
The 0.66 eV Match: The energy required to jump that first “hump” corresponds almost exactly to the 0.66 eV (1878 nm)control barrier I discussed above.
The Router’s Role: Without an “optical router” like melanin to capture and down-convert cosmic flux, the environment can’t provide the precise, coherent push needed to get over that peak. The energy just stays as “noise” (heat), and the reaction stalls.
19. The Proton Gradient as the “Dark Sector Bleed”
The red line labeled “Proton gradient” is the “shortcut.”
The 1878 nm Harmonic: As I discussed above, CCO (and its Archean precursors) uses the 1878 nm harmonic to resonant-pump protons.
The “Work”: This proton gradient provides the electromotive force that lowers the activation energy of the first step. It transforms a “difficult” geochemical reaction into a “spontaneous” biological one.
The Lorentz Connection: The “Lorentz-steered flux” provides the initial high-energy particles that melanin uses to “charge” this proton battery.
20. Methanogenesis: The First “Solid-State” Output: The future substrate of Archeae
The graph ends at CH4 (Methane).
Archean Feedback Loop: As life (the Methanogenic path) successfully turned CO2 and H2 into Methane, it created the very Methane Haze (the “Titan Analog”) that I discussed earlier.
The Information Engine: This haze then buffered more cosmic flux, providing a feedback loop that stabilized the 0.66 eV environment. This is exactly what I meant by “Information from Flux”, the system creates the conditions for its own expansion by reducing the uncertainty of the environment.
21. Why “Synthetic Gasoline” Matters
The note on the image, “The first steps of this reaction are difficult – try making synthetic gasoline”, is a nod to industrial catalysis.
The Catalyst: In industry, we use massive heat or transition metal catalysts (like Iron or Nickel) to force this reaction.
The Biology: Life does it at “room temperature” by using Ferredoxins ([Fe-S] clusters) as the solid-state wires and the proton gradient as the power source.
My Thesis: Life is a self-assembling refinery that uses the Resistance Principle to bleed the dark sector’s pressure into the production of organic matter. Water is the key power source, not ATP.

This image is the “proof of concept” for my Solid-State Metabolism. It shows that the difference between a “dead” planet (the Geochemical path) and a “living” one (the Methanogenic path) is simply the ability to establish a Proton Gradient, which is the very “Quantum Battery” that our 0.66 eV melanin router was built to charge. What does this imply?

THERE SHOULD NEVER BE ANOTHER QUESTION WHY THIS SLIDE IS ACCURATE FROM ANY PATRON OR MEMBER.

Centralized medicine labels decentralized, biophysical models as “quackery” because they operate outside the chemical-industrial paradigm. Modern centralized medicine is built on the “medical casino” model: it treats the body as a closed chemical system where symptoms are fixed with exogenous molecules (drugs).
Decentralized medicine is perceived as “crazy” because it identifies the human Lagrangian as a solid-state, optical process. It shifts the focus from “caloric intake” to “flux harvesting,” which cannot be patented or sold as a pill.
The Source of the “Quackery” Label
The medical establishment relies on Newtonian biochemistry, which ignores the quantum foundations I’ve outlined:
The Isotope Blind Spot: Centralized science ignores the Kinetic Isotope Effect (KIE). It treats all hydrogen as equal, failing to see that deuterium (“the grit”)increases the inertial drag of the CSF and ruins the 3rd and 4th “bends” of proteins.
The pH Misunderstanding: It views a 1.5 gastric pH solely as a digestive tool, missing its role as the primary deuterium exhaust for the brain’s quantum hardware.
The “Inhibition” Fallacy: It sees Nitric Oxide (NO) at the CCO site as an inhibitor (poison) because it stops ATP production. It cannot perceive the “Impedance Adjustment” that allows the system to switch to high-capacitance “Harvesting Mode.”
Why My “Wicked Game” with my tribe is Necessary
The world is “on fire” because the modern environment is a frequency mismatch.
The 1878 nm (0.66 eV) Gap: Indoor life lacks the SWIR harmonic required for resonant harmonic pumping of protons. Without this, the S8-Ferredoxin tunnel clogs, and the “teleportation” of protons stops.
The Lorentz Collapse: Without UV-A to flip the NO switch, the body stays in “High-Resistance” mode. It burns food exclusively, creating the “Biological Fry-Out” (heteroplasmy) that leads to addiction and chronic disease.
People call this quackery because it implies that the front lobe’s “advancements”(technology, artificial light, processed food) are actually biological traps that decouple us from the Lorentz-steered flux of the sun.
The Bottom Line is this in this series of blogs
Decentralized medicine isn’t about affirmations; it’s about dielectric constant restoration. It recognizes that Nature is the Savior because only the 1878 nm harmonic and UV-A can unlock the CCO gate and “un-weight” the human system from the entropic drag of deuterium.
By pointing back to the “soil illuminated by the sun,” I’m exposing the medical casino’s reliance on “heavy” hardwarethat has lost its optical coherence due to the artificial light it is forcing mammals to live under today. That is the asteroid behind all our ills.
SUMMARY
I’ve identified that Biochemistry as the “shadow” of Solid-State Physics which really powers life. ATP is not what sustains us, it is the water that CCO makes that does (above). We are essentially “dark sector lightning rods” that have learned to use the resistance of our own carbon-based hardware to weave the fabric of life.
Melanin is our Ancient Optical Router for Cosmic organization before codes. Before codes there was flux and melanin controls that flux of electrons and protons. I’ve identify melanin/water battery in modern cells as the primary “Dark Matter Sensor”. Because melanin has a complex chiral and chaotic carbon-ring structure, it can interface with these high end radiation from cosmic signaling. The proof it can happen is occuring in Chernobyl right now. This implies that melanin and ferrodoxin were likely the first useful molecules on the planet before life made in the primoridal soup by self organizing principles.
Food webs only come from the photosynthetic web. Photosynthesis (Visible Light/CO2) is a “luxury” light that built a biochemical metabolism that required a shielded planet by ozone. This is the one your doctor learns about in medical school. Radiosynthesis is the “atavistic” engine designed for the raw, high-entropy chaos of the cosmos that is stepped down by hydrated melanin. This is the one that is present inside Chernobyl right now regenerating all the life in Ukraine that the nuclear plant put at risk in 1986.
Every time the sky turns grey, your body doesn’t “shut down metabolically”, does it? No, it pivots back to the Archean Earth’s power grid for the wisdom that was built by 4.6 billion year old Archean photonics.
My thesis has shown that the human is a fantastic energetic machine that tells 4D time and it carries the entire history of the Earth’s relationship with the Sun in its melanized circuitry. We are not a “product” of evolution; we are the persistence of the flow from our star to planet and we are an electrical resistance to that flow that has allowed the information of light to unfurl into life. This is what evolution is all about. Darwin and creation missed large portions of this recipe by design. Melanin allows us to convert 10,000 times more energy than the sun every second and the Rockefeller Dynasty has built a world since 1911 that makes sure no doctor ever learns this truth.