
Most people who have watched my Vermont 2017 youtube have come away astonished at the insights I put together there using the science published in the last 100 years. This blog might be more shocking to those who fully grasped the complexity of the physiology of the retina. I briefly mentioned in the Vermont 2017 talk the role of Muller cells and RBC’s in the retina. What I did not then, is that both of those cells in the human retina operate using a non-pathologic Warburg metabolism. That talk gave an extreme level of quantum biology and was done to show you how obesity begins in the eye. Obesity is extended when the signals in the skin are also usurped by a blue light environment. I made a prediction in my own hacks 13 years ago that if the CT protocol and Leptin Rx were true, that a melanopsin like photoreceptor had to be present in the skin, its arterioles, and subcutaneous fat mass to explain obesity in humans fully. Little did I know in 2017 right after my Vermont talk that paper got published and my educated guess was correct.
I did not mention this in Vermont 2017, because the next step in your quantum education links directly to what happens in the skin. This is why I was provocative with the audience there last year and I suggested to the Dave Hollenbeck that if he wants to see some more amazing quantum biology he should have me back in Vermont in 2018 to talk about how the skin operates with the peripheral clocks in every organ in humans. It turns out he took me up on the challenge. I went there on June 2, 2018 and unleashed my ideas on the topic. So my members and Patrons get the early heads up on what innovations are coming in this talk in Vermont. Seeing the details on the slides I have made, in person, might just blow your mind.
The teleological explanation for the presence of the Warburg effect in the mammalian retina is simple to understand once you have a basic idea of what Krebs bicycle really is. When proteins synthesis is massively upregulated by ubiquitin, the cell cannot rely on the steady removal of anions from the TCA or urea cycle to fuel biosynthesis because of slowed down kinetics of either cycle. This creates a logjam for the cell with respect to the cell cycle. The collateral effects are felt in the matrix, with respect to hydrogen movements, because of the need for of H+ to run the ATPase to power both cycles. So what does a cell do when redox drops and the TCA and urea cycle spin rates are lowered for any reason?
It relies on the older evolutionary pathways, glycolysis with the help of H+ to transfer information over energy to get the job done. Each C-N bond in proteins costs 5 ATP. This is very costly on an energy basis. Information costs are even greater. This means biosynthesis is an energy hog for the cell. Does the retina use the TCA or urea cycle exclusively for biosynthesis? No, they do not. This should surprise people considering the metabolic rate of the retina. So how does this process occur in the retina? The retina is a tissue with a high oxygen demand in the back of the eye but a poor supply in the front of the eye.
The anterior chamber of the eye has to rely on getting oxygen from diffusion through the cornea from the air because the cornea and lens must remain transparent for vision and no blood vessels can impede the passage of light. If the cells of the eye used TCA intermediates exclusively for biosynthesis, considering this oxygen issue, it would create a chronic proliferative state in areas of the eye and this would impede its function. This defeats the purpose and the physiologic requirements of the central retinal pathways, pituitary, and hypothalamus. All of these places in the brain that are light relay stations are quite small in size. The reason these parts of the brain are small in stature is that they deal almost exclusively in information quanta processing over a growth and proliferation state.
This implies that the retina must run two different metabolisms to get the job done in vision. Is this true? Yes, it is. It turns out the adult mammalian retina is non-proliferative because of how it is built to interact with light, it shares similar biosynthesis requirements to neoplastic tissue because of the prodigious turnover of the opsin proteins in the disc membranes of the photoreceptor outer segments. This explains why certain cells of the retina use the Pasteur effect. You had a blog on that effect recently here at Patreon. Each mammalian rod outer segment consists of a stack of ∼1500 distinct discs enclosed by the plasma membrane. Approximately 60% of the dry weight of the disc membrane is protein, of which, opsin comprises 90% of the protein content. Remember all opsins are “loosely” covalently bound to Vitamin A to operate in humans. Hence, the retinal-bound opsin, rhodopsin forms a large structural component of the rod disc membrane. These components are even larger in the outer retina where melanopsin controls the quantum release of melatonin release from the pineal after 4 hours of dark. This is the major quantum mechanism that controls mitophagy and autophagy signaling.
All G-coupled receptors are linked to the circadian mechanism in humans. Rhodopsin is a G protein-coupled receptor comprising 348 amino acids, with a rich glycine and serine component. This is the very same serine and glycine amino acid components that link them to the efficiency in the PPP (and glycolysis). The PPP is the major way we regenerate the major reductive moiety called NADPH you learned about years ago in the EMF 4 blog post.
Tissues with poor oxygneation cannot use the TCA or urea cycle. This is critical in tissues that cannot use the TCA or urea cycles because this is a backdoor way for the cell to augment informational quanta transfer using H+ pulled from the serine cleavage system. RBC’s, Muller cells and every single embryonal stem cell uses glycolysis and the PPP by nature’s design. None of them have high oxygen contents. This keeps a lid on their growth rates. There is NOTHING pathologic about these cells that have to rely on gluconeogenesis. The key point in understanding is that when cells use gluconeogenesis for biosynthesis exclusively they MUST use glycolysis and the PPP for biosynthesis. What determines their use? The amount of oxygen tensions in the tissues is deterministic for the pathway a cell uses. Glycolysis and the PPP are dominant when oxygen tensions are low!
This is where you may remember David Sinclair paper about pseudohypoxia, NAD+ and aging I mentioned in December of 2013 on the blog. Sinclair was talking about cells losing the ability to use the TCA and urea cycle as they age so they all seemed to have a common phenotype of having low NAD+ and pseudohypoxia. He called it a cardinal feature of aging and disease. It turns out he miss read the tea leaves. As cells age, they become less able to use the TCA and urea cycle because autophagy loses its efficiency, due to a loss of redox power. When autophagy becomes faulty, sleep is poor as a marker, and as a result, a cell loses control of atoms of deuterium buried in the PUFA in the mitochondrial membranes. You heard about this mechanism in the May 2018 webinar. This is a huge big deal folks. Why? What are the collateral effects of this mechanism? Is this how cells make and create the ELF-UV light they use for a mitotic division? Yes. Is this the basis of the research of Fritz Popp work on ELF-UV light. Yes.

Richard Young did the seminal experiments on the retina. The deuterium critics have no clue how this complex dance operates because they do not understand how information is transferred via the quantum spin state of protons and electrons to Orbital Angular Momentum (OAM). Moreover, they have no idea what the OAM is used to do in your cells or blood. The next few ideas link my Vermont 2017 talk to the one I gave on the skin last week. This is a story of how deuterium does things people never anticipated to help mitochondria and cells self-regulate their own behavior under the power of the Auger effect (UV light). UV light is critical in controlling ECT flow in mitochondria several ways as this series has shown.
Now you do and I am showing you how it links to the physiology that controls melatonin creation and releases in the mammalian retina. In fact, this quantum mechanism is so powerful that Mother Nature decided to use it kingdom-wide in our clade and on every surface of their body plans.
The rhodopsin turnover rate in the human retina parallels the degree of aerobic glycolysis found in many different species in mammals. The rhodopsin turnover rate matches the Vitamin A and D cycles in humans almost perfectly. I showed a slide of this in Vermont. This is why unsupplemented Vitamin D levels are great proxies for defects in Vitamin A cycles in the brain and retina. This makes your unsupplemented Vitamin D3 level a great proxy for what is really going on in your skin and eyes mitochondrial matrix. The next part of the puzzle is not well known.
Photoreceptors have a relatively low rate of turnover in lower vertebrates, and it is temperature dependent (another clue to the H+/D effect). Recall that higher temps favor H+ bonding in aqueous environments versus D bonding in water. As temperature increases, the non-pathologic Warburg effect becomes temperature-dependent. In mammals the story is exactly the opposite. The rate of turnover in the mammalian retina is astounding and this is why mammals became warm-blooded. They used heat release from their mitochondria to precisely control the flow of H+/D in the matrix and cytosol to control growth and metabolism of the retina just by using light and temperature and oxygen concentrations in the eye. This is the basis of the CT 4 and 6 blogs written over a decade now. All of these reasons are why I made the educated guess years ago that melanopsin had to be present in the skin/fat of mammals. In December of 2017, I was proven correct.
Since melanopsin is present in the eye, arteries of the skin and fat mass this becomes a critical point in understanding how the skin becomes the gatekeeper of deuterium flows from the blood plasma versus the tissue. Remember phototaxis is well known in the leaves of plants. Most third graders know that leaves grow toward light and can change their angle of inclination of growth toward sunlight based upon the type of sunlight present. Very few people realize that the RBC’s in mammals do the very same thing as leaves. This maybe why aneurysms form in arteries because RBC function differently when they are out of sunlight. Arteries should have laminar flow in the center of a vessel and why blood flow is more turbulent towards the artery wall. Sunlight causes the entire arteriole to migrate toward the sunlight of UVA light. Sunlight can directly affect the RBC’s in the center of your blood vessels to act just as leaves do to collect IRA and UVA light as a form of animal phototaxis.
Recall UV spectrum in sunlight heats mammalian skin up the most significantly. IR light in sunlight is considered cold light energy on a relative basis but it is the initial light of the sun that builds the EZ in tissues and our blood. Remember, via chemistry’s laws that heating favors H+ in hydrogen bonding over deuterium bonding, so organisms can use their body heat as a fractionation method to control growth in tissues. This helps you understand further why mammals evolved warm bloodedness. Heat is a powerful physical tool in controlling the flow of different isotopes of hydrogen in your cells. It turns out UV light raises the temperature more than any other light waves in sunlight.

Having the innate ability to uncoupling your protons gradients helps keeps deuterium in the blood plasma because blood moves so rapidly, it can diffuse its heat like a radiator, but tissues do not have the same ability. This relative difference is all that is needed to control where isotopes of hydrogen should be found. When deuterium is trapped in the arterioles of the skin is can be pressurized by sunlight in the UV range and the chiral heat effect to pinch deuterium in the blood vessels coming to the surface, while at deeper levels H+ accumulation is favored in our tissues. Compression of H+ or deuterium allows the body to make a continuous UV spectrum from the isoform of hydrogen trapped in both tissues.
The UV spectrum is not equivalent because of the chiral heat effect. In the blood, the spectrum of light extends deep into the UVC range, and this frequency matches the optical absorption spectrum of the EZ in the blood. Recall this is built by the water and IRA light initially from the sun. When this phenomenon occurs in the blood plasma, the EZ can grow further to absorb light rays into the UVC range to further energize blood plasma to carry more light energy and information in the ophthalmic artery distribution of the retina. The flow of information is more important in tissues that cannot use the TCA or urea cycle. This helps the areas of the retina that have poor blood flow. This circulatory pinch of hydrogen allows deuterium and hydrogen to become a creator of full spectrum UV spectrum light creators via an unusual nuclear effect specific to hydrogen on the periodic table. This photic phenomenon extends the size of the EZ in the blood and vascular space to become something more than just a battery or capacitor as Pollack has shown in his studies on water.
The NIR light addition to blood allows water in our bodies to transform into an optical resonator or a mirror. This mirror can then be used to optical resonator to create tiny lasers at our skin surface or in our ophthalmic artery which feeds the retina. That UV light created by this mechanism is then used to regenerate melatonin, dopamine, and build many more substances in quantum thermodynamic fashion. This system is amazing when you see how the quantum system is organized to support the semiconductive circuits of the central retinal pathways I laid out in last years Vermont talk. How it links to the skin is also quite remarkable. You might want to watch that Vermont video 2018 when it goes live here on Patreon.
We can see the wisdom of information quanta by nature when we carefully examine the anatomy of the photoreceptors in our eyes. I showed some of Dr. Wunsch slides in Vermont in 2017 but I could not get into this level of complexity with that audience because they were not facile with the science and they were not mitochondriacs who understood what Krebs bicycle was. I am hoping to change that this year. You have to understand how quantum thermodynamics differs from classical thermodynamics to understand the process fully.
In the retina information from light is the key resource that determines the anatomy. The organization of the retinal cells involves the highly compartmentalized cellular configurations, with the confinement of mitochondria to the inner segment, absent from the outer segment. The dense aggregation of mitochondria in the ellipsoid region of the inner segment reflects the considerable reliance of this portion on oxidative energy from the TCA/urea cycle. Evidence that supports these ideas was the finding of high concentration of malate dehydrogenase (an enzyme involved in the TCA cycle) in the primate photoreceptors inner segments. The malate result revealed a level 30 times higher than the outer segment of the photoreceptor. Malate needs fumerase to be completely free of deuterium to add water to it to support the functioning of the inner retina. This tells the wise person nature had to build a light-based control system to control the flow of H+/D if any retina was to operate with sunlight. I began to put all the pieces together 15 years ago. Now you can see information processing in the eye and skin really begins with H+ and deuterium fractionation occurring immediately in the surfaces of the retina and skin. This helps explain why nature really built the retina in the way she did. For those of you who don’t know the retina appears to build inside out. This has confused biologists for ages because none of them understand light well. The same idea is true in the skin’s physiology.

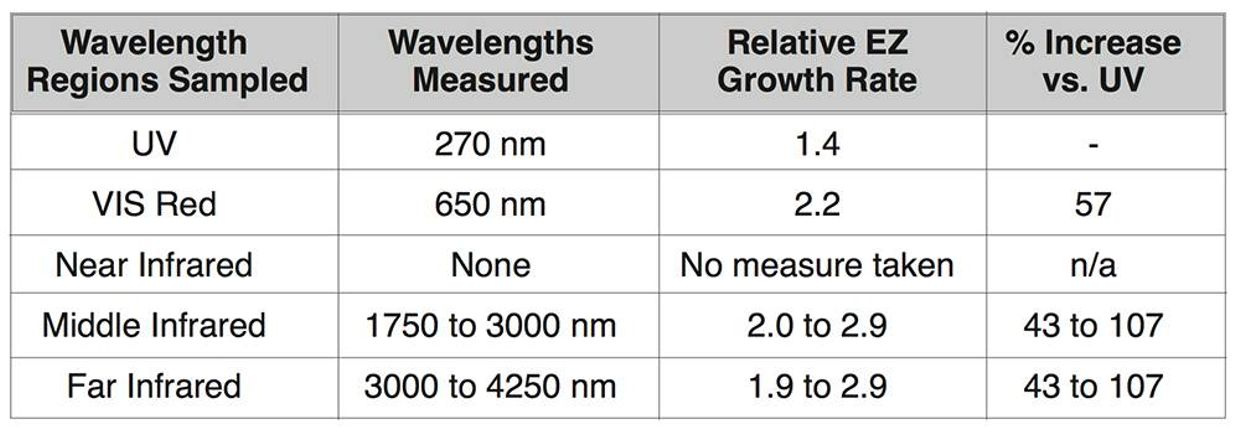
Since this was true in the retina, I knew that it had to share metabolism with several pathways that worked with different oxygen levels. I hypothesized that we should see data that lactate would be higher in areas where the PPP and glycolysis are used in the retina. I believed strongly in 2003 the same thing had to be true in the skin because of their shared embryology. Do we have any evidence of this? Yes we do. Contrary to conventional wisdom, the measured fractions of lactate dehydrogenase (LDH) used in glycolysis was significantly LOWER in the inner segment where the cones are located as compared with the outer segments of the retina where melanopsin and Vitamin A are used. This tells you cone vision needs lowered oxygen tensions because the light they work with is less powered than the blue light that melanopsin works with.
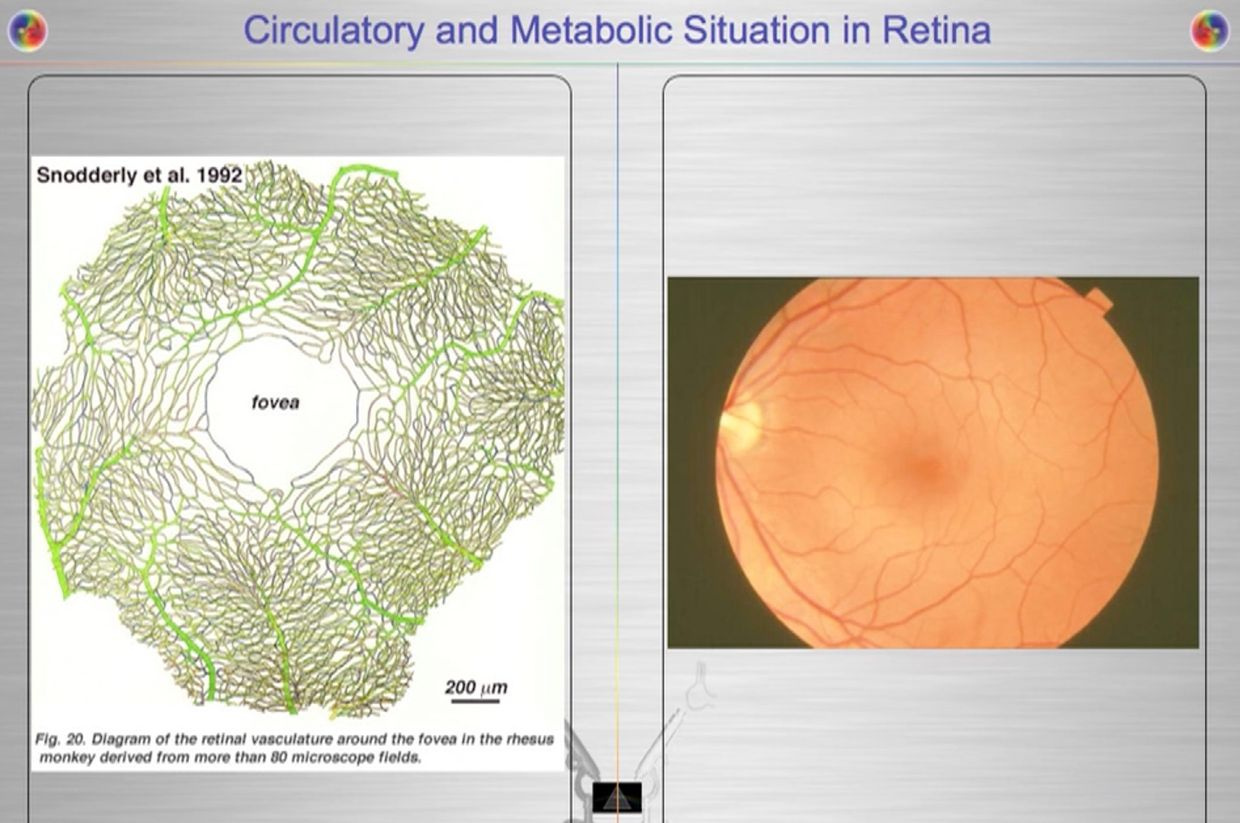
This is why the fovea/macula has a POOR blood supply. I should those pictures last year in Vermont (above). How does the retina accomplish these gymnastics? It did exactly what the RBC did in evolution. It eliminated mitochondria from the cone cells while restricting blood flow to make sure no TCA/urea cycle metabolism could occur there. Anyplace mitochondria are, deuterium must be rare, and vice versa. The exclusion of mitochondria from the photoreceptor outer segment necessitates its reliance on glycolysis/PPP for cellular energy production, while still permitting information transfer from sunlight via protons with the H+ isoform in the retina and into the small structures of the brain mentioned above. There is a deep lesson here for the mitochondriac about how and why the retina where built as they were to work with hydrogen to make UV light inside our body to drive photonic programming of physiology. This is how information quantum is created and moved in the central retinal pathways to affect your brain and CSF pathways to build the world wide web in your head called your mind.
CITES:
https://onlinelibrary.wiley.com/doi/pdf/10.1111/ceo.12462
https://elifesciences.org/articles/29217
Dr. Jack Kruse Vermont 2017 Youtube talk
Dr. Jack Kruse Vermont 2018 talk