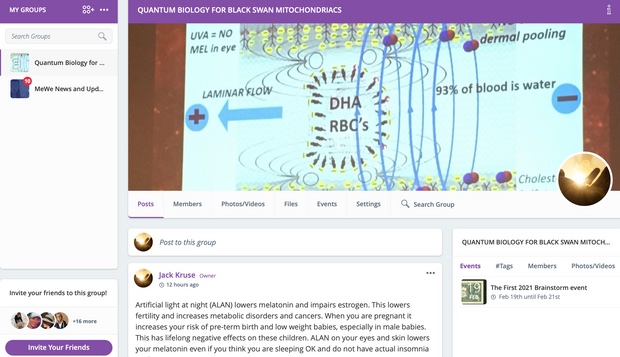
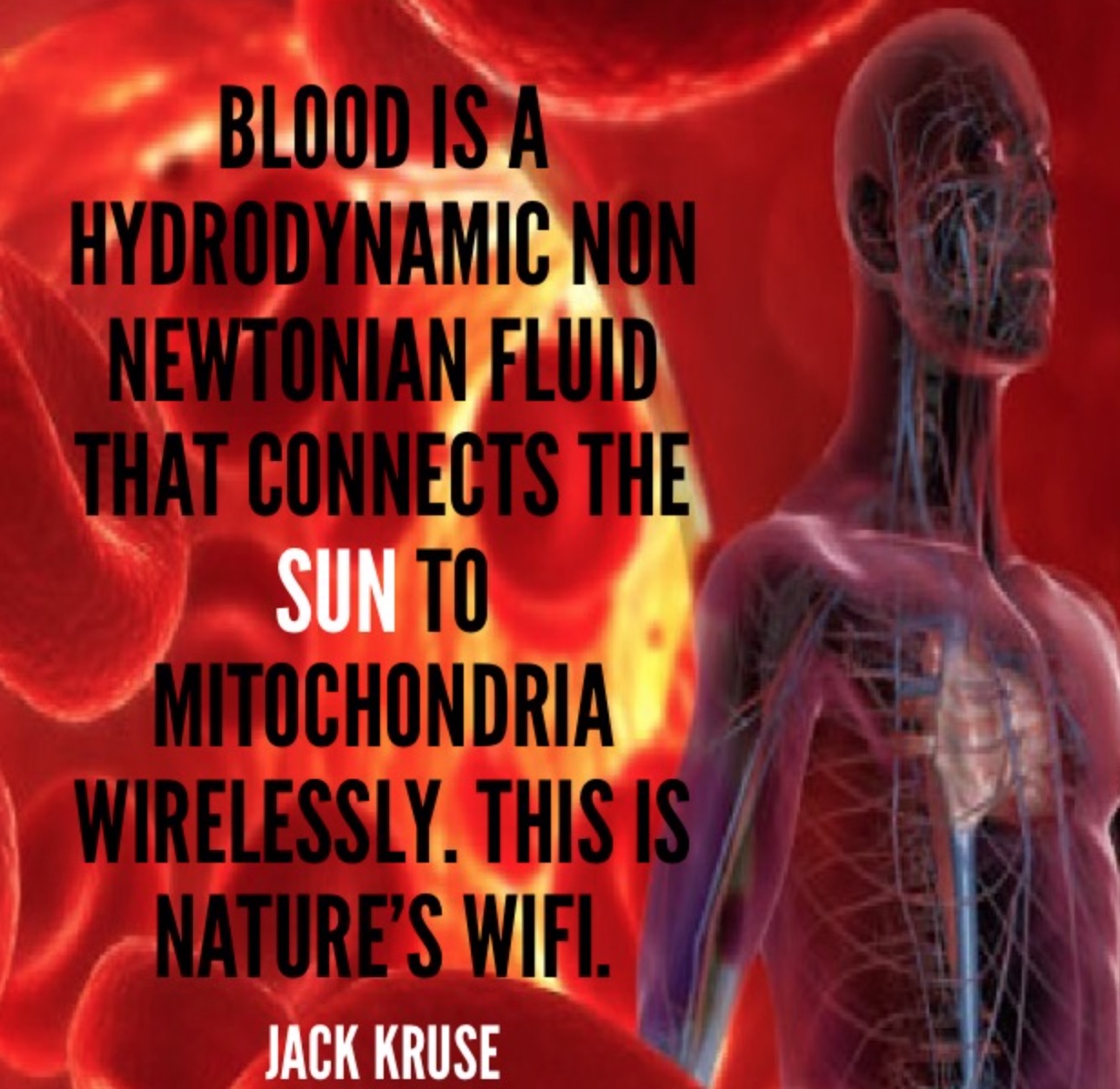
Why take an Rx blood thinner when the sun does it for free? Does this explain why darker skin people get COVID more, and get severe clotting issues with COVID, and die more frequently when they live at high latitudes and in cities where nnEMF and blue toxicity dominate? Can you explain this Uncle Jack? Look at the picture above and realize what is going on in blood when your skin is irradiated by terrestrial sunlight. Do you think the same waves are made in the blood when you are under fake light?
Do plants grow the same in the sun or with grow lights? Might this problem show up in us too?
Our modern lifestyle dehydrates us chronically and causes us to lose the ability to create Vitamin D and use magnesium in a proper thermodynamic way in cells. Both of these problems lead to clotting and a lack of immunity. It also is the reason why Cancer shows up in people who avoid the sun because of idiotic public health beliefs that the sun is toxic. This is our greatest risk factor for skin cancer in my opinion. Australia is the classic example of this modern risk factor today. Anyone who is struggling to get their Vitamin D levels higher likely has a co-morbid magnesium deficiency due to a lack of water from poor mitochondrial function at cytochrome C oxidase. People forget that one of the main by-products of mitochondrial energy transformation is the production is water at cytochrome C oxidase.
Many of these people dehydrate their colony of mitochondria when they live in an environment loaded with non-native EMF and blue light. What is the first thing that happens in the mitochondria when this occurs? It reduces the amount of melatonin the mitochondria make. This impairs both autophagy and apoptosis. When this happens neither change program is efficient at removing defective engines and disease present. That is Why COVID, CLOTTING, and CANCERS ARE all linked to a lack of sun and too much tech exposure.
Magnesium in a cell is a hydrophilic element on the periodic table, and without water, we lose intracellular Magnesium (Mg). 56 enzymes in mitochondria use Mg2+ as a cofactor. It turns out making melatonin is a Magnesium and water-dependent process. Making Vitamin D is also water and Mg dependent. There are 3 metabolic transactions from Cholesterol (Cholecalciferol) that occur under the surface of the skin where our “storage version” of Vitamin D (Calcidiol) is made. This 25(OH) version of D3 gets transformed in our kidneys and liver to the “active version” of Vitamin D (Calcitriol or 1,2,5 (OH) in the liver. All 3 reactions require Mg2+ as a cofactor. Mg, however, needs the mitochondria to make cell water in the cell to work properly. If the mitochondria do not make water, taking an Mg supplement is a waste of time and resources. It is physiologically impossible to have a 25(OH) blood test (“Storage-D”) to be less than 35 ng/dl and a Magnesium RBC level to be above 6.5mg/dL because of the negative feedback loops tied to calcium levels in our blood!
As a result, this is why melatonin levels are off in these patients. With time this ruins their sleep because melatonin forms another coupled cycle with cortisol and adenosine. This is why Magnesium and Vitamin D deficits walk together in mitochondrial damaged patients. This is why COVID people get the side effects they get. Some kids with COVID have enough redox power to avoid clotting but they get chronic fatigue instead. It will not change until their redox power is returned by changing the environment they are in. If this is allowed to persist chronically it is only a matter of time that clotting will occur or that will develop another chronic mitochondrial disease, like cancers. I think this is why Oz is loaded with melanoma.
If you want success in life, become a problem solver for your patients. Not a fixer.
Magnesium is also a co-factor for telomerase that controls our telomere lengths. When you marry this with a low melatonin level you begin to see why cancers will come as these people age. It is also the reason why dark people get the worse outcomes from COVID and why they COVID to begin with. Taking a vaccine won’t solve this problem but getting in the sun will. I also think people who use a ton of sunscreen are at risk for clotting, COVID, and chronic fatigue. What else exacerbates this linkage? A modern diet with undiagnosed high O6/O3 ratio due to seed oil usage with the use of carbohydrates out of season which also causes dehydration. One mole of fat creates 100-110 nmol of water. One mole of carbs produces 55 nmol of water in the mitochondria. Carbohydrates also contain more deuterium, and deuterium main function in the body when it gets into the mitochondrial matrix is to decrease the piezoelectric ability of the mitochondrial via the increased kinetic isotope effect. IT makes the mitochondria act more like a diamond, and less like quartz. A diamond carbon lattice is not compressible, while the SiO2 lattice of quartz is. This is why quartz is piezoelectric and why a diamond aren’t. This is why Nature only creates carbs in strong light cycles when the sun can offset the lack of metabolic water production from the sun in mitochondria at cytochrome c oxidase.
In medical school, I learned that Vitamin D was a hormone that was important in bone metabolism and that is where it really ended until I began to read about Vitamin D3 15 years ago. In all that published research, it uncovered what it really does for our cells. The major effect is that it modulates the immune system as its receptor is found on T helper cells (regulator cells), the same cells that are destroyed by COVID spike protein or how HIV infection wipes out in AIDS victims. So if your Vitamin D levels are low, FOR ANY REASON, they turn off the epigenetic switches on RNA/DNA/mtDNA genes that turn on the proper functioning of immune system protector cells to keeps us free of diseases from viruses, bacteria, mold, and fungal infections. In fact, since I got my level checked 15 years ago, I have not been seriously ill. The sun provides what we need. No mRNA can do this. When you give a person with low redox and a low D level an mRNA vaccine we should expect new variants of COVID that are more lethal. It appears our Public Health experts in the centralized medical systems did not expect this. My clients, in my decentralized practice, however, did.
SUMMARY
Artificial light creates a positive charge in most of the things in a cell. Galactic cosmic radiation also has positive nuclei and charge. These positive charges were accelerated in the shockwaves of supernovae throughout the galaxy and they randomly enter our solar system.
Redox power from the sun, manifests in living systems is all about optimizing the net negative charge in cells.
Its proxy is found in the charge carrying ability of water (dielectric potential) made in mitochondria which control intracellular endogenous glutathione levels. All things that cause inflammation carry a net positive charge. This is why pH matters in the care of COVID patients in a hospital. It predicts who is really in trouble.
If you need a clotting drug, you are, BY DEFINITION already blue light toxic, nnEMF overdosed, and get far too little sun on all parts of your body.
Creators are hard-driving, focused, dominant, independent risk-takers. Who are the emerging players on top of the Monopoly board in DeFi?
DeFi update Feb 5, 2021:
DeFi is becoming a cash flow machine. These protocols are generating annualized earnings (est.): Uniswap = $140M (60% of swaps) Aave = $100M (interest & flash loans) Sushi = $90M (15% of swaps) Maker = $45M (seniorage-based credit swap lines)
Most protocols are <2 years old. Ponder that.
We’re on the ground floor of incredible change that is on the way in DeFi!
The current DeFi narrative depends very heavily on the future of Ethereum. It is highly questionable now if it can ever evolve into a network, that has e.g. identity, governance, privacy, and scalability issues completely resolved in a manner, that’s required from a true financial services network. My bet is on something, where those four things are solved first.
I think paying attention to the volume of the coin of dollar business they do will give us clues who will crush it on ETH 2.0 monopoly board I spoke about earlier in this series. It will also tell us ETHER 2.0 is the real ground to rebuild the new monopoly board under DeFi financial services.
Genius isn’t necessarily coming up with everything from scratch, it’s knowing how to take some existing ideas and perceive them in a radically different way to come to a new way of thinking that can unlock value in something that’s very, very new. Your ability to critically think you must engage your imagination. This blog is about my synthesis of where the axis of value is being built on the Monopoly board. The true sign of wisdom is not the knowledge you acquire, it is how the data you feed into your brain is filtered through yourimagination to get to a place where you can see the results of innovation before anyone else does.
My top 5 crypto right now Feb 2021:
1. Bitcoin
2. Aave
3. Uma
4. YFI
5. ETH
I think 2, 3, 4 are the first pieces being added to the Monopoly board. I think they are lots on the board with foundations being poured now. It is early…..but what they do is all about how many of us are getting yield on our crypto accounts.
WHAT IS Aave?
For example, Aave is best described as a decentralized system of lending pools.
Users deposit funds they wish to lend on ETH2.0, which are then collected into a pool. Borrowers may then draw from those pools when they take out a loan. These tokens can be traded or transferred as a lender wishes.
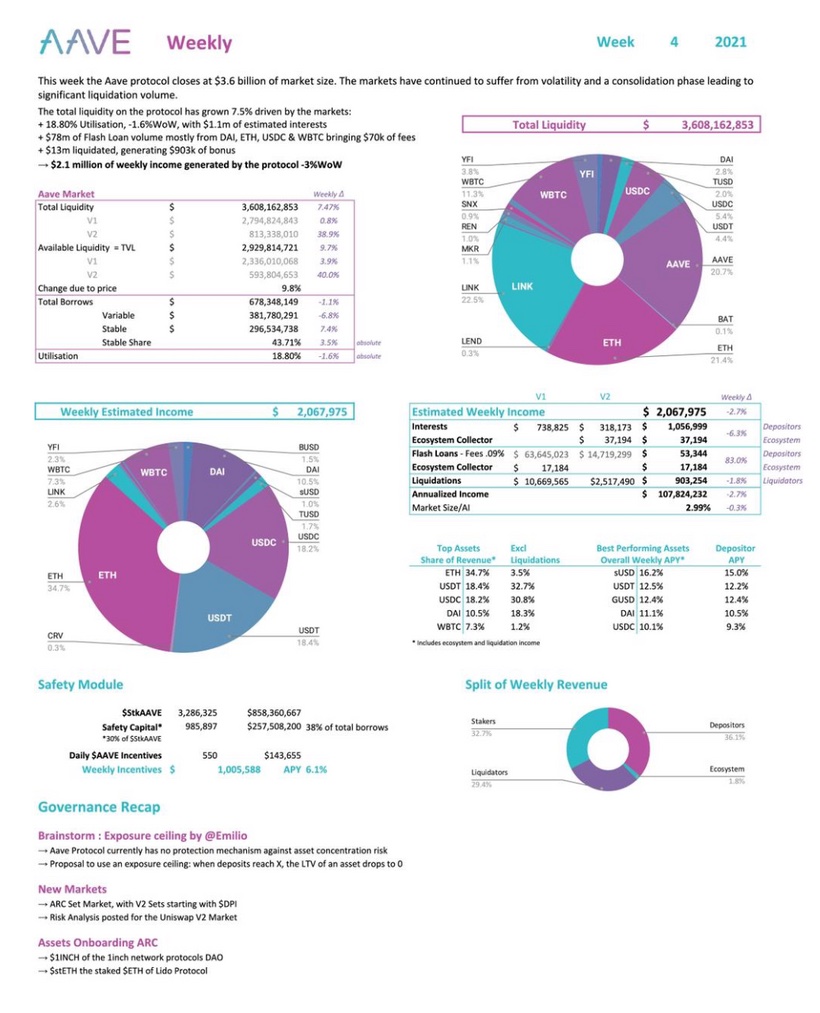
To make it clear regarding revenues of ($aave): Estimated weekly Income : USD 2,067,975 Weekly incentives : USD 143,655 Yield APY : 6,1% Price to Sales : 67,5 x Volume/Market Cap : 19,58% DeFi is growing faster that anyone can imagine.
Why does AAVE have intrinsic value?
As the Patreon blog says, ETH 2.0 is not decentralized but Aave protocol is. That is why I think it becomes a key property on the monopoly board. AAVE plays a central role in the management of the Aave software, allowing users to vote on changes to its rules and policies.
Aave was released in mid-2020 and has went from 80-430 since I first bought it when I was in my last few days in Mexico in Dec 2020.
AAVE owner are able to vote on interest rates for certain loans and aspects of how deposits are managed. This makes it interesting to me. Interest is how banks operate. Aave is a replacement for bankers in the DeFi space. Aside from this utility, AAVE derives value from its finite supply, and the fact that it uses revenue from fees to buy AAVE and remove the cryptocurrency from circulation. This makes it decentralized.
Currently, 80% of fees collected by the Aave system are used to burn AAVE. THIS TELLS ME SUPPLY IS SHRINKING SO THE PRICE HAS TO GO HIGHER. I believe it might go higher than any other coin in 2021. The remainder is used to pay lenders in their space. The constant burning of AAVE is expected to reduce its supply, thus driving up the price of the token if demand remains constant. So far it has worked as I thought.
The 4 elements I have cited here (identity, governance, privacy, & scalability) are essential elements for a truly decentralized financial system. They are four legs of the DeFi table, if you don’t have one the system will eventually be out of balance and mean revert to the intrinsic value of the labor put into the code. For me, this is why I am not yet sold long term on ETH 2.0 as a chronic winner. I do think what is being built on it, may far surpass its value. I want to discuss how I see the “early axis of this monopoly board” is being built today in front of eyes.
Many don’t understand how projects using the Ethereum 2.0 protocol equates to higher Ethereum token prices, so here is how it happens. As the DeFi projects continue to go up like Aave, UMA, UNI, YFI it means ETH has to be used up to complete the transactions.
The way ETH 2.0 goes up is Gas. ETHER is the gas that makes the DeFi projects operate. So for a more accurate answer, I suggest doing some homegrown research.
My understanding is that actions taken on the Ethereum blockchain have operating costs per transaction. Right now early in the ETH 2.0 game gas is expensive. If you have to pay for the gas you hate this increased cost. If you do not, and just own ETHER you’ll love this because consumption of ETHER because it cuts supply and this eventually will drive the price. The problem is there is too much ETHER in the market now to drive huge price gains, but this is going to change over time and makes ETHER look like a great play ten years out in the way I view the market. Since ETH 2.0 just started in the last 6 months, gas is giving people sticker shock, but if you when you realize there are 37 billion money transactions already in DeFi projects you’ll begin to understand why this situation is something to pay attention to and understand clearly.
When you want to compute something on the blockchain, you have to pay for that privilege. That action can take the form of sending tokens from one ledger to another, or it can be the execution of a smart contract. That cost replaces the subtracted middlemen. This payment happens in something called Gas = ETHER. And Gas comes from ETH tokens. So any project on the ETH blockchain needs ETH to run. It burns Gas. Gas comes from ETH tokens. Every project on the ETH blockchain creates a demand for Gas, which creates a demand for ETH tokens. And this increase in demand leads to a higher token price. This is driving ETHER price movement now. This begs the question, if ETHER IS going to move, then what crypto assets are fueling this momentum?
UMA = the VIX
The Chicago Board of exchange (Cboe) Volatility Index, or VIX, is a real-time market index representing the market’s expectations for volatility over the coming 30 days. Investors use the VIX to measure the level of risk, fear, or stress in the market when making investment decisions.
For those of you who do not know what UMA does……it is the VIX of crypto. The VIX of crypto right now just got off the ground in the DeFi space. It controls 60% of DeFi swaps in this space as of today.
Professional traders use volatility data to make informed decisions and be on the right side of trade. Therefore, the CBOE Volatility Index, known as the VIX, is popular with equity traders.
Bitcoin is known for its high volatility and Universal Market Access (UMA) plans to offer traders an opportunity to trade volatility in a decentralized way. I think this coin has huge potential. For that, the protocol plans to launch a uVol-BTC product that will be settled at the end of the month. Traders could use the token to speculate on the increase or decrease in volatility, or hedge their Bitcoin positions they do not want to sell during sharp corrections. Today on Feb 5, 2021 options expired on BTC and the price is moving. I expect the derivative market for BTC to skyrocket over the next ten years. If the product turns out to be successful, a uVOL-ETH token is also planned in the future.
Other than the regular synthetic tokens that track the price of an asset, traders keep looking for new opportunities that do not give institutions a direct edge. In order to cater to this demand, UMA plans to launch a new synthetic token called ‘uSTONKS’ that will be based on the ten most commented stocks on the r/wallstreetbets Reddit forum. This is another reason I am bullish on UMA. The new uSTONKS token offers retail traders a unique way to benefit from the movement of the most popularly discussed stocks on the forum.
Early discussions are also in progress to create a “Big Mac Synth” based on the index invented by The Economist back in 1986. Launching innovative tokens attracts new traders. New traders = Metcalfe effect for the entire DeFI space. Another reason that could have added to the short-term demand for UMA was the negative 15.5% annualized yield for depositing Ether as collateral to borrow USDC.
UMA has soared from an intraday low at $11.234 on Feb.2 to an intraday high at $43.998 today, a 291% gain within three days. I began buying it in December 2020.
The first batch of tokens to be listed is uSTONKS-MAR21 with 10 stocks: GEM, AMC, NOK, BB, SLV, TSLA, PLTR, SNDL, AAPL, and SPCE. UMA is launching the index in collaboration with YamFinance which is one of the most popular farming DeFi projects.
UMA price could surge even higher according to various metrics
It’s important to note that despite the recent price surge, the number of whales holding between 100,000 and 1,000,000 coins has increased by seven since January 28. Similarly, other large holders with 10,000 to 100,000 coins have also joined the network in the past week which indicates investors believe the digital asset has the potential to rise higher.
WHAT ELSE?
Aave is a digital finance app on ETH 2.0 that is building a digital trust score which is analogous to an Equifax credit score in the finance world. Aave is a money market that is attempting to establish the time value of digital assets by paying interest on the movement of things with value on Blockchains. It does not have to be just money or coins. It can also be a property that is tokenized. It is the world’s first true digital lending market in my opinion.
For example in Sept 2020, UniSwap exceeded the crypto exchange CoinBase in terms of exchange of value as a Decentralized market. That got my attention last year and I decided to look at all the players that were just beginning on the ETHER chain. How Uniswap made money was more shocking to me.
On Coinbase exchange the exchange owner gets the transaction fees from trading volumes. They also had the risk of building and funding the exchange. What Uniswap was, as if you owned their token you would share in the transaction fees directly with coin ownership because the Dapp called Uniswap would cut out the middle which in this case was Coinbase. Once I understood this I began building a position for all these companies collecting fees in swaps. Uni, UMA, AAve, Market, YFI are right now my favorite ways to play this DeFi trend. Owning them is a bet on future commerce in the DeFI space. I will say not all of them will succeed long term. Some will go to ZERO. Be aware of this. Another way I think about this speculative play is that one of these companies might become the Amazon or Goldman Sachs of the DeFi space. That excites me and I think this space might offer more upside than BTC will. But it filled with execution risks.
The person who holds the coins is getting paid as transactions go up by the value of the coin going up. In reality, it is like being paid a dividend yield. It is a new way to own a bank without the equity risk. This group of coins is actually building the Yield curve for the DeFI world. In the centralized world, these companies are worth a fortune. Right now you can buy these coins very cheaply.
The problem is knowing which one is winning the battle of supremacy. I follow the money flows to make a decision and I add to those positions. I am fully aware that 90% of these names might go to zero but I have faith that one will become the 300-pound gorilla of the DeFi space.
The biggest development I think is each one of the companies makes money every time someone buys BTC and decided to get a yield on it. Many people. like, me, are doing this. If you view BTC as pristine collateral that is growing 200% why in the hell wouldn’t the market want another 6-10% of BTC yield as the BTC sits in a cold storage vault run by YFI, as you wait for BTC to go to 1 million a coin? The way I think about it, if I want this so will a lot of others. Therefore, I want to own some of these coins.
This is a market that has no need for venture capital when you really understand it. The market is self-funding. That means this space is going to eliminate the VC in the analog market. How much is that worth?
In 2019, North America had the highest valueof VC deal funding globally, with approximately 113 billion U.S. dollars in financing. Right now these companies market cap on crypto exchanges is only 15 billion dollars. That tells me they are worth a ton more.
Can this space be used to create easy free profit via arbitrage? What is arbitrage?
Have you noticed BTC or ETH have different priceson different exchanges at the same time when you’re buying? This is how you arbitrage the price.
Arbitrage is already profitable in DeFi.
Arbitrage traders are an essential component of the Uniswap ecosystem. These are traders that specialize in finding price discrepancies across multiple exchanges and use them to secure a profit. For example, if bitcoin was trading on Kraken for $35,500 and Binance at $35,450, you could buy bitcoin on Binance and sell it on Kraken to secure an easy profit. If done with large volumes it’s possible to bank a considerable profit with relatively low risk. Uniswap and Aave do this now.
How does this actually work?
Getting started with Uniswap is relatively straightforward, however, you will need to make sure you already have an ERC-20 supported wallet setup such as MetaMask, WalletConnect, Coinbase wallet, Portis, or Fortmatic.
Once you have one of those wallets, you need to add ether to it in order to trade on Uniswap and pay for gas – this is what Ethereum transaction fees are called. Gas payments vary in price depending on how many people are using the network. Most ERC-20 compatible wallet services give you three choices when making a payment over the Ethereum blockchain: slow, medium, or fast.
Slow is the cheapest option, fast is the most expensive and medium is somewhere in between. This determines how quickly your transaction is processed by Ethereum network miners. Right now the gas fees on ETH 2.0 are high. That is great for ETHER owners because it uses ETH supply up to lower the supply and this makes ETHER less centralized the more it is used by DeFi application. So in essence I am saying these decentralized apps and tokens are what is driving ETHER real value right now. It is not ETHER itself. It is a giant feedback loop being built in the DeFi space. The more Aave, UNI, UMA, and MAKER money they move, the better it is for ETHER price.
If my thesis is correct over the next 4 years we should see ETHER price moves move away from BTC moves and it should happen after these DeFi apps keep driving capital across the DeFi ecosystem.
Now that I understand this space well I think each one of these DeFi apps is fully composable onto many other businesses that will get built on ETH 2.0. I personally think banks are going to favor ETH 2.0 because they believe it is a centralized platform they can control. DeFi apps are going to make ETH 2.0 more decentralized. So I see Aave, UMA, YFI, Maker, as the main competition for banks in the next few years. They have a huge advantage because the yields they are paying on crypto exchanges cannot be matched in conventional banking. Banks however, know when interest rates go negative money will flow out of their bank into something else. They will adopt blockchain and hope their clients use their payment rails into the world of crypto. I think they are nuts to think this way. DeFi is the best way to unbank and this is why I love these coins over the next 5 years. These coins compete with one another, but unlike banks, they fit together like lego blocks in the DeFi system and there is no longer a financial problem of Bank of America connecting with JP Morgan in this financial world. I think that is a really valuable that legacy banks won’t understand how to adapt too. I think Avanti bank in Wyoming is going to disrupt them in a big way. If you do not follow Caitlin Long you should. She is a Wall St. veteran who is now going fully digital with Avanti. She has deep ties to Kraken and to Senate banking committee via Senator Lummis.
My Take
Senator Cynthia Lummis has been assigned to the Senate Banking Committee.
Lummis has been vocally pro-digital assets in both her candidacy and since taking office.
Lummis said she plans on forming a Financial Innovation Caucus to educate her fellow senators on emerging financial technologies. She was key in changing Wyoming laws for the benefit of Kraken and Avanti who are both based in Wyoming. She is going to be a big player in the Senate for Bitcoin.
The Office of the Comptroller of the Currency (OCC) granted another crypto-focused firm conditional approval to operate as a national bank.
Protego will join Anchorage as a banking editor with approval from the federal government, according to a new announcement from the U.S. banking regulator.
Protego & Anchorage aim to serve institutional clients interested in digital assets. The Seattle-based firm plans on being a “vertically integrated and fully regulated bank built on blockchain.” Now, as a nationally chartered trust bank, it can hold, trade, lend and issue digital assets to clients. These will be its four primary services, according to the bank.
However, Protego is still in the organizational phase of development, according to the OCC’s announcement. It will have 18 months to meet the terms of its conditional approval. It will be able to convert to a national trust bank when it begins to operate. Currently, it’s a Washington State-chartered trust company.
I am not sure which lego block DeFi app is the most valuable yet, so I decided to spread my bets across a bunch of them.
This video lays out why the leaders of ETH 2.0 cannot be trusted.
Tolerance of ambiguity is a necessary condition for creativity for wealth and health creation.
Perfect money has never existed, of course until now, gold is the closest thing to it. The shiny metal might be a pet rock or a barbarous relic to some, but it has remained the most stable asset over the last 5000 years, granting stability every time monetary collapse ensues; the best candidate to replace whatever the elites claim is backing the fiat monetary system is now underway. Bitcoin is a solution outside of the governments power. I believe it will be a large part of the solution of the coming financial tsunami in the next 5-9 years in this Fourth Turning.
In the early 1800s, during the first of George Friedman’s U.S. socioeconomic cycles, the greed of the Second Bank of the United States’ stockholders created an economic depression. To enrich themselves, they issued paper currency with no backing, prompting rampant speculation, ultimately leading to a crash. (Pic of Jackson from New Orleans)
The 1837 Panic ensued as President Andrew Jackson, a sound money advocate, had responded to the speculation by backing the U.S dollar with gold and silver to purge economic deformities from the previous cycle. The economy remained somewhat stable until America fought a civil war thirty years later. Abraham Lincoln’s government committed the same economic suicide as the shareholders of the now-defunct Second Bank of the United States. That bank issued vast sums of paper currency to finance the Civil War by printing money— plus speculation on railways — drove America into yet another depression, which led to the Coinage Act of 1873 where President Ulysses S. Grant enacted the de facto gold standard to suppress malinvestment and inflation. This is where the gold standard began to end the hyperinflation of money printing.
This period in U.S. history is what American’s should expect when the next crisis unfolds today in front of eyes, only this time with a modern twist: Cryptocurrencies will be used in some way to replace gold. The next major collapse will coincide with both the mass adoption of crypto and the resurgence of commodity money in some fashion.
As paper fiat money rapidly devalues something else the elites will use will reflate the value of money. This will lead the resurgence of commodity money around the world. I expect it to be fully digital. The COVID pandemic has already begun to warn people to stop using fiat and used debit cards and credit cards and I view this as step one of the government warning the public something new is coming. They will use the pandemic to ease into a new monetary reality. I believe this is why the pandemic really was orchestrated as it has been. It provided a cover for the Treasury for us to avoid another civil or revolutionary war. I believe this is why the elites in Washington DC have made the January 6th reactions of the people so noteworthy in their response. Furthermore, I believe that is why permanent fences are being installed in the Capitol. If they did this without the events of January 6th they’d have no plausible answer why they did this.
Detractors like me, are cautioning you against the symbolism of permanent fencing surrounding the building. A government run by criminals needs protection from the “We The People”. This is precisely what Snowden and Assange have warned us about. Now we can see why lawmakers have asked for this. Lawmakers say keeping a barrier around “The People’s House” cuts against a message of transparency and accountability to their constituents. This is a farce for the Great Reset s, real agenda, monetary debasement.
Permanent fencing sends the wrong message to the nation and the world, by transforming our democracy from one that is accessible and of the people to one that is exclusive and fearful of its citizens because Congress is a cabal.
Crypto suits an increasingly technological world in which we desire speed, security, and privacy. It is programmable money and a form of digital gold. Using precious metals as a currency — trading in grams, and transporting freight loads between parties like in the California Gold Rush of 1848 — is a crazy proposal for a modern medium of exchange. Transacting with a crypto wallet beats transacting with a gold debit card, hands down.
Crypto, however, isn’t perfect money either. At best, it’s a currency without commodity backing. It is backed by scarcity and the promise of elimination of a central banks controlling arm. It removes politicians and banks control. Though we live in a society where the elites have replaced gold with debt, this doesn’t mean we must continue their mission to destroy money’s intrinsic value. We have the choice to opt out and unbank ourselves before their plan takes effect. As history shows, we need a gold-backed currency to reverse the sickness that fiat currencies have inflicted on society, and to have any chance of preventing elites from siphoning off wealth for themselves via monetary manipulation.
Only when the great reset occurs — where the fiat system relinquishes, and money regains its value, meaning, and purpose — will we know whether this new monetary paradigm will be governed by the people for the people, or by the same type of elites who always finds a way to exploit the system. It’s obvious which outcome will create a better, fairer, juster world, but it’s also clear, based on history, that the power will end up in the hands of a more autocratic, digitally empowered elite. Is this the moment when the people finally rise up and take charge? We’re about to find out. Is BTC packing your parachute, because it is packing mine. I have lost all faith in the Fed and US Treasury.
I believe the day Decentralized medicine was born was the day the Nobel Prize in medicine was awarded in 2017.
What is cool about bringing old science to new eyes by putting windex on the glass eyes of medicine?
Your certificates, degrees, status and wallet do not matter. How you treat your equals and people under neath your direction matter most. Doctors and patients are not speaking the same language to one another because they both suffer from the same ecologic deficits these days, while have a different educational paradigm of beliefs = They are both chronically disconnected from circadian biology and nature. Doctors were schooled in a centralized system which is their algorithm of understanding. Their customers are not biased in this way. They get this information for free, and this makes them the product being sold to the industrial healthcare complex.
If I can get one in hundred physicians and patients to see this perspective, then I am doing OK. This new project I am involved with on Quantum Health TV aims to inflate these ideas for physicians to build a new decentralized medical system.
Medical school 25 years ago could be considered an algorithm controled by a curriculum. This curriculum allowed maximum human intervention. It did not allow machines to take data and then decide for patients.
Algorithms: For those in non-procedural medical fields it’s our thought process we offer our patients. And rigorous logical medical thought is becoming scarce, I fear because they are importing our knowledge base into algorithms and the output of algorithms continues to morph because machines continue their algorithm variance. There is no control. We are now training medicine residents that will easily be replaced by midlevel practitioners who haven’t gone to medical school but know how to apply algorithms in recipe format.
But everyone forgets the outcome of algorithms continue to evolve long past the day they were created.
What we fail to realize is that contemporary technology permits decentralization, it also permits centralization via algorithms when they go unchecked by human critical thinking. It depends on how you use the algorithmic machine learning and how you let machine learning use you.
Decentralized medicine is mitochondrial based and involves significant physician input. The key issue for medical students and young physicians, is can they hope to capture a job in DeMed platform in the future? Can they do this without going through 10 years of debt creation and a centralized educational format taught in training? I have found they can if the are connected to the digital monetary network to power their careers as early as possible. This creates freedom to untether from the centralized hospital system.
Can you imagine living in a world where executing a care plan or operation, buying an original idea to solve cancer, or accepting payment from a patient can be done without needing trust? We might be in position to just trust the code of a blockchain? No third party, like a hospital or insurance company is required?
What is obvious is hard for those with a bias to accept. There are two kinds of people in this world. Those who think the government is doing things in their best interests and those who think critically, to make our world a better place.
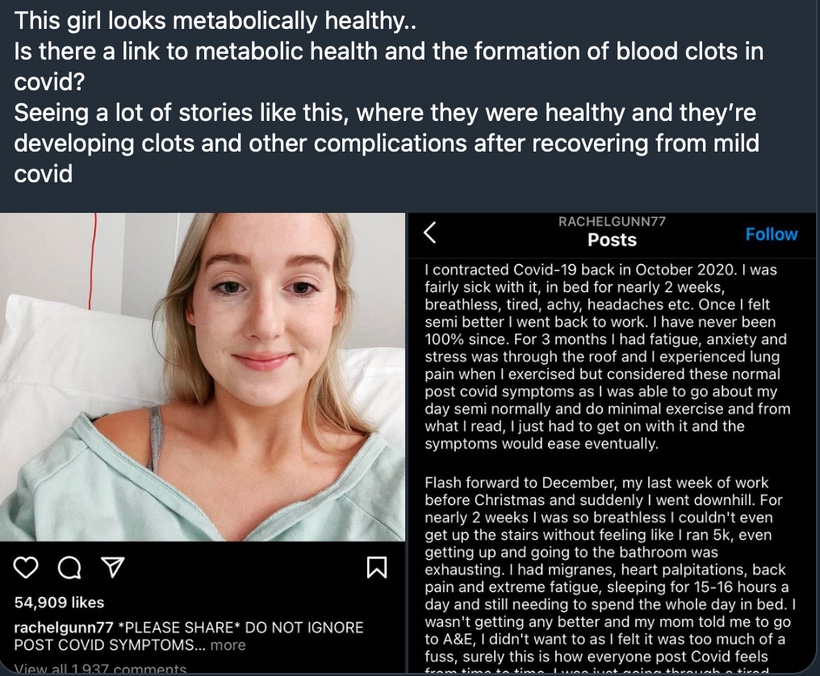
Pictures like this below make sense to a decentralized doc, but not to a centralized one. Are you really healthy if your blood clots when you get a virus? Centralize docs have no idea how the lack of a proper Coulomb force can lead to clotting in COVID. Coulomb force, also called electrostatic force or Coulomb interaction, is the attraction or repulsion of particles or objects because of their electric charge. When people clot platelets charge has changed for some reason found in our light environment. Electric charges cause electromagnetic force fields around them. This causes things to do the things they do.
Coulomb forces must be balanced after daytime and this occurs at night when light is absent. This is why light at night is so bad for us, and why a centralize doc has no ability to figure out why this girl clots her blood after becoming exposed to a weak virus. Does this tell you something deeper about humanity now? Why are humans dying from a weak virus now? A decentralized doc gets it…………do you?
We see the world as what we carry in our own hearts. When others see mistakes, I observe a lesson.
They see a shadow, I observe the light that casts it.
They see the night, I observe the moon and its effect from the Earth’s tilt and how it varies the Coulomb force. They see a corporation, I observe people who make it thrive and die. They see millennials as a problem, I observe our bright future. They see success, I observe the coming journey. They see me as a stranger. I observe them, as my future friend and cohort in mito-hacking for a better world.
^^^^^^^WHY IS THIS AN ACCURATE DECENTRALIZED VIEW POINT?
Melatonin is created by the Coulomb forces in AM sunlight. Melatonin levels vary within tissues because of how much light tissues get. As a result, it appears melatonin is a guardian of the Mitochondrial genome (37 genes). Out of the 37 gene, 13 of them are specific to energy-transforming mitochondrial genes. Those 13 genes only code for the proteins that tunnel electrons and spit protons out of the mouth of cytochrome proteins. The mitochondrial genome undergoes 3 times as many genomic mutations as the nuclear genome by design. Melatonin levels are critical in monitoring mitochondrial behavior and light energy transformation by controlling how energy is being transformed from light to create physiologic power.
The major function that melatonin helps smooth-out in mitochondria are
Energy metabolism and flux via control of electron and proton tunneling.
Redox balance within the mitochondria = GSSH
Ion homeostasis = electrostatic controls via Coulomb forces that vary via light
The signaling of cell death and mitophagy = % heteroplasmy in a mitochondrion = the amount of light energy transformed to physiologic power.
This picture defines the centralized system. In the decentralized system There are over ten metrics to evaluate thyroid function. Centralized testing looks at two. If those two are within normal limits, “you don’t have a problem”. You get handed a Rx for an antidepressant and synthroid.
The only way to fix the the medical system is to build a decentralized one where no one on the treatment side of care has ultimate control but the patient. The relationship between doctor patient must be repaired. Three mechanisms in which decentralization impacts on health system equity, efficiency and resilience are:
1. ‘Voting with feet’ reflects how decentralization either exacerbates or assuages the existing patterns of inequities in the distribution of people, resources and outcomes in a jurisdiction.
2. ‘Close to ground’reflects how bringing governance closer to the people allows for use of local initiative, information, feedback, input and control.
3. ‘Watching the watchers’ reflects mutual accountability and support relations between multiple centers of governance which are multiplied by decentralization, involving governments at different levels and also community health committees and health boards
Where others see nothing or impossible strings………I observe decentralized answers. We have to cut the umbilical cord physicians have with centralized systems to get to the level of understanding how environmental light controls reality we observe. That won’t be easy because most physicians are now paid by that system. Physicians must plug into the BTC monetary network before they cut their umbilical cord. The more physicians become employed by fiat payors the more subject patients will be to corporate medical algorithms. These computer generated prescriptions will determine your health outcome. You and your doctor lose their power to help reverse the disease.
The current centralized payment structure reinforces the focus on disease by rewarding procedures, not cheaper interventions like prevention or care coordination. They never pay for therapies that hold the promise of disease reversals.
In the 1950s, 60s and into the 70s, primary care physicians were well respected members of the community and helped patients navigate and coordinate more specialized care. Children of this era remember having a family doctor who would attend to all of their family’s medical needs.
In the 1980s and 1990s when I was in medical school I was affected by how the system incentivized care, as the cost of healthcare became burdensome for corporate and government payers, the dynamic changed. The federal government and the insurance companies created a structure of procedures and payment rates for each. Procedures based on higher levels of training and technology received higher fees. The Feds and insurers tried to push down prices of procedures, but at the same time they rewarded advances in medical knowledge and technology, and the result was highly trained specialists were well paid for performing sophisticated procedures, and family doctors were squeezed. This trend was a driver in my own decision tree back then on what career path I should choose.
IMPLICATIONS
As a result of the centralized system, treatment decisions are influenced by money, not necessarily what is best for the patient. The procedure-based payment structure rewards doctors for doing more, even when it might be better to do nothing. I know this first hand. This is still true in 2021 for complex spinal surgery.
Our healthcare delivery system spends more than 700 billion of its 2.3 trillion in annual health spending on medical care that does nothing to improve a patient’s health…seven hundred billion dollars every year. And, most alarmingly all that ineffective treatment and harmful care represents one-third of tests, treatments and procedures that physicians perform.
Our current centralized reimbursement system and our cultural values serve up a ready answer to physician uncertainty as to what tests and treatments to order for their patients: more is better. When evidence is incomplete or conflicting about when to use a particular procedure, surgery or diagnostic test…some physicians will treat more aggressively, especially if piecework reimbursement rewards that.
The implication: Unfortunately, only about 20 percent of clinical procedures have solid scientific evidence to back them up. This means in many case physicians are flying blind and under great economic pressure to do more, even when it doesn’t necessarily serve the patient’s needs. With an arsenal of government-approved treatments available that are paid for in a centralized manner, there is great temptation to do “something,” even when it might be better to simply watch and wait. I saw that unfold in my own practice for 25 years.
With money flowing to specialists, primary care doctors were forced to see more and more patients and had less time to spend with any one patient. Unpaid services such as preventive care and care coordination quickly went out the window. With the lure of greater prestige and earning potential, medical students funnelled into specialties, creating a deficit of primary care doctors. Today, it isn’t unusual for patients to be shuttled from one specialist to another with no one looking at the bigger picture of the patient’s well-being. This process has gotten larger over my career. Patient outcomes have gotten worse.
Most of you know my specialty is neurosurgery, and within it, I subspecialize in spinal reconstruction. You would think, from this position I have obtained I would support centralization. The position has allowed me to see the problems clearly. This position showed me that centralized power in medicine is an impediment to health of people. This is why began to speak out against this centralized model of care 15 years ago. My personal perspective is the generalist in science always is the most valuable specialist. They know more so they connect more dots to lead to a new understanding to get health back.
In centralized medicine patients’ preferences, goals and values are marginalized.
When the patient is reduced to a vehicle for disease, the doctor becomes the most important person in the healthcare process. This may work fine when medical decisions are straightforward. In acute diseases they are. In chronic disease management they are not.
When there is ambiguity, a patient’s preferences, goals and values are essential in choosing the right course of action. This is a strength in a decentralized paradigm. The current centralized culture of medicine, in addition to the economic incentives mentioned above, doesn’t encourage this kind of two-way communication.
The discussion about norms and values inherent in every clinical judgment and decision seem to shift from the doctor’s consultation room to the conference room of the doctor’s professional association. If intentionally or unconsciously physicians do not want to negotiate with their patient about the usefulness of certain interventions, they can refer to the opinion of their professional association that is codified in guidelines and protocols instead, thereby shifting the responsibility for clinical decisions from a personal decision to a professional group decision. That is a deep centralization problem.
All too often, the patient buys into the mindset that decisions about their health are best left to evidence based paradigms of associations. Patients become passive recipients, rather than active participants in their own care. This may have been acceptable when the aim of medicine was simply to keep people alive, but chronic conditions, in particular, require the patient to play a larger role in managing their own health.
Adding fuel to the fire is a rising tide of chronic illness. Simultaneously, the Baby Boom generation, which represents a quarter of the U.S. population, is beginning to hit retirement age. This is a period of life when we become more vulnerable to illness and chronic conditions tend to accumulate.
As we’ve seen, the disease-based and doctor-centered medicine that brought us so far in the 20th century isn’t well equipped to mitigate and manage this growing tide of chronic disease. A new decentralized paradigm is needed that treats the whole patient and establishes a more balanced relationship between doctors and their patients.
Today, in medicine progress is measured by the speed at which algorithms can destroy the conditions that sustain life. Those narratives are pushed on social media as evidence based by doctors and researchers employed by the centralized system. The evidence of the fidelity of those “evidence based ideas” are found in the abysmal results of our chronic disease epidemics that the centralized system.
Our centralized healthcare system in the 21st century focuses on treating diseases, not people. The medical knowledge we gained in the 20th century had very narrow goals: stop people from dying. It was focused on treating short bouts of illness caused by a specific disease often localized to a particular organ or organ system. However, the CDC estimates that over half of adults in the U.S. suffer from one or more chronic diseasesthat cannot be cured, only managed via machine learning algorithms. The costs of treating these diseases now represents 75 percent of the $2.3 trillion in U.S. annual healthcare spending.
The centralized system fosters these beliefs………..Do you?
I know I do not.
DECENTRALIZED MEDICINE IS A SOLUTION
We can use algorithms as an aid to the systems of our society, like pilots use autopilot or how we use cruise control in our cars, but we must never let them run our society completely on their own – the day we do, will be the day we fall.
The right thing isn’t always real obvious to the rest of the world. Sometimes the right thing for one person is the wrong thing for someone else. And everything, in retrospect, is obvious to those who use forethought. How you think makes you great. 99.9% of humans use hindsight to dictate their actions. Hindset in medicine is dangerous. We have let the industrial healthcare complex take over decision making and no one is questioning the problems with it. Some of us train to observe the unseen first. We learn to ignore what the eye sees and focus on what the brain observes. This is how the decentralized mind sees problem solving.
Like most things in life, everything is great— until it isn’t. Circadian destruction comes from bluelight and nnEMF and this has created our chronic disease epidemic reality.
DeMed is the ‘counterintuitive” antidote for nnEMF ecocide caused by technocracy. With ecocide being the irreversible destruction of our natural environment by electropollution.
The irony of modern society is that we are now the best-informed society that has ever walked this planet, but yet we also carry the greatest risk of dying from our own ignorance. Technology allows for escapism from nature. Being divorced from Nature is where all diseases begin. But technology can be harnessed to change medicine just as easily.
This is why good people can make tragic errors for people who follow them. Think employed docs and patients.
Case in point below.
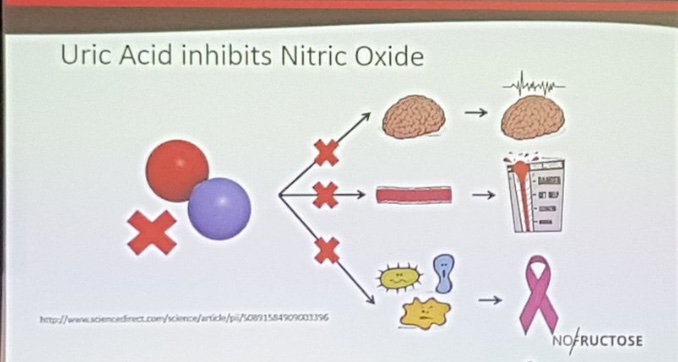
How does a half-truth about food lead to a full lie in reality? The LCHF folks say this: Increase fructose load increases uric acid which inhibits the production of nitric oxide constricting blood vessels to the brain. Is this true?
Recent literature shows that uric acid is actually protective to damaged human brain tissue. So how come the LCHF physicans got it all wrong?
Did they see the whole truth or just the truth they understood from their centralized education?
The reality of the entire picture: Fructose shows up in foods grown by nature that only grow in strong photosynthetic environments naturally which have full-spectrum sunlight. So fructose is never designed to be present without strong solar UV/IR exposure. Nitric oxide is also made by short wavelength UV light, so who cares if fructose inhibits Nitric oxide? Nature always couples them…………..
The only people who do care are people eating an LCHF diet who live indoors way too much. They never get nitric oxide from their skin because they are rarely in it to experience it so their physiology is tuned the way nature designed. This is also why they get fat. The subcutaneous fat in humans responds metabolically and hormonally to sunlight or the lack thereof. Sunlight affects the Coulomb charge of our bodies. It just so happens sunlight increases Nitric oxide production to offset the loss in foods with fructose. So it is really a lack of sunlight and not the foodstuff that is the real problem. These physicians never get to this level of understanding because of their centralized education that was paid for by Big Pharma and the Industrial healthcare complex.
Big Pharma is not interested in cures, just creating new customers. This is why blue check MDs are dangerous for the public. They never ask the right questions for you because of the algorithms that were uploaded to their minds in training. Critical thinking has atrophied in physicians who have been trained using corporate algorithms.
Medicine is now a tech industry where clinical nuance is dead and the centralized algorithms of the health lobbist rules public health.
Decentralized medicine allows the doctor to examine the algorithm and critique with by holding the coding outcome up to Natures rules to limit the risk of disease.
Why do I love writting my ideas down for you? I enjoy self-publishing & sending researchers and clinicians copies of my blogs. What these blogs represent are rejection letters for the paradigms beliefs. When these clinicians and reearchers read my work, they respond, ‘Who is this guy?’
And I’m like, ‘the end of your industry.”
Decentralized medicine is a virus worth spreading.
SUMMARY
Blockchains allow humans to connect weird things together
Many people even today do not understand how we can create a software of silicon to hold a greater value than gold. That is BTC in a nutshell.
How can blockchain technology bring value to medicine?
Communications is at the heart of medical e-commerce and community. Telemedicine has exploded on the scene because of COVID. Soon physicians will be paid in Satoshi minute by minute for their value. Apps like Strike will facilitate this transition. Banks like Avanti will fuel our practices. They are building on and off ramps we will need to build the practices our patients need.
The global eCommerce market poised to reach $135 billion by 2023. The introduction of blockchain technology in eCommerce helps build a global decentralized economy. Medicine has to be part of this. It is projected that within a decade crypto could be a mainstream alternative to cash and cards.
Physicians need to know the consumer’s appetite for crypto payments is influenced by good publicity, ease of use and convenience. They’ll never get this experience as an employee in a hospital. For merchants and retailers, crypto payments will likely be an add-on to existing systems. One thing we can agree on is the payments industry is dated and ripe for disruption in medicine. Physicians should embrace this chaos and not run from it.
Physicians have to become capable of accepting and processing cryptocurrencies. It will foster “a marriage between inventory management and exchange platform.” This decreases physicians needs for centralized systems of support. Blockchain technology is capable of handling virtually the entire eCommerce experience for physicians who go out on their own. This includes product searches, supply chain tracking, loyalty programs, product reviews, data security, and post-sale customer care.
Blockchain technology facilitates verification and trust while eliminating the need for a middleman. In the case of medicine, the application reduces the dependence on Big Pharma and researchers who perform useless science and useless papers, and electronic medical records. The blockchain ledger will help us keep an accurate database of a work’s provenance on the blockchain. This data can be monetized by patients and physicians and not by Google or your hospital.
DeMed physicians will be the innovators of this paradigm shift. The forward thinkers and risk-takers are necessary in every technological advancement and revolution. Many of these ideas will fail and billions of dollars will be lost and dreams shattered. From these attempts, the best of the best will rise, and change how we live, work, learn and play. Failure is always the first step to success. I know. I have been failing at this paradigm shift for 15 years but I refuse to give up on this idea. Success in building this new practice is stumbling from failure to failure with no loss of enthusiasm
I believe the world is ready now for this idea.
The current GME stock squeeze situation and DeMed are linked. Do you see the link yet?
The moral of the story of GME is that retail Reddit investors, for better or for worse, finally learned how to weaponize options and the short squeeze operation using leverage. lt is likely going to lead to the utter destruction of the current centralized markets beginning today, thus, setting the stage for globalized decentralized markets that are fueled by BTC monetary network.
Doctors and nurses need to find how to optimize the BTC monetary network to power their successes while limiting their own destruction at the hands of corporate algorithms, medical schools, residencies, and hospitals. So when your monetary network is inflated, no person or employer can force you to vaccinate yourself or do something to a patient that breaks Nature’s laws for mitochondrial function.
DeMed defends chaos. Decentralized systems are the quintessential patrons of simplicity. They allow complexity to rise to a level at which it is sustainable, and no higher.
American federalism was the embodiment of political tolerance and decentralization – the expression of the liberal conviction that society can manage itself and needs no central plan.
American medicine before the AMA was the embodiment of political tolerance of ideology with no central recipes. Where patients can manage their diseases when reintroduced into Nature.
The purpose of a centralized medical system or any other system, is not to ensure stability in people, but to exploit people in the society. It is time to change the paradigm.
Physicians and nurse interested in transition planning can join me for brainstorming sessions on how to get the logistics right at kruseatdestin.com. DeMed amateurs talk tactics. Professional DeMed users talk logistics.
Earlier this month, Britain’s Financial Conduct Authority warned crypto traders that they “should be prepared to lose all their money” because there are so many risks involved in Bitcoin.
Furthermore, Janet Yellen, President Biden’s pick to head the Treasury, laid down the hammer when she voiced worries last week that decentralized, anonymous cryptocurrencies could be used by criminals — provoking fears of a regulatory clampdown under the Biden administration.
“I think many are used, at least in a transactions sense, mainly for illicit financing, and I think we really need to examine ways in which we can curtail their use and make sure that money laundering does not occur through these channels,” Yellen said Tuesday.
This sounds like very scary stuff for Bitcoin.
Except it’s not.
What just happened in the centralized fiat markets with GME and AMC is a far bigger risk to investors. The actions are directly linked to M1 and M2 money printing too. Someone should tell Yellen this. Maybe she is deaf because she got 810,000 dollars to speak from the Citadel Hedge Fund. the same fund that just defrauded Robinhood users? She got a lot more money from Wall Street by reports out today in the media.
Ironic……..that as soon as Yellen tried to scare Bitcoiner’s the shit storm just hit her own windshield. How does Wall Street cheat? It is not with crypto………they cheat with fiat money instruments.
The shorts, used fiat regulations of leverage to sell the stock of GameStop 140% short. Did you hear one person on social media or TV complaining about Wall Street hedge funds with short positions trying to crush GameStop claim? Not one. They were 140% short because they used fiat leverage to do it!” How did they do it, Ms. Yellen?
They used cheap fiat money from the Fed printing press to gain leverage using rehypothecation.
GME was rehypothecated, which is a fancy way of saying that brokerages lent out shares of GME that didn’t exist. They basically created stock when there was no stock. Hence why the short interest was above 100%. It was at 140% yesterday and today it is still at 127%. This is what happens when you trust a third party. Bitcoin doesn’t require a trusted third party because it is DECENTRALIZED!
Central banks can print money far longer than ‘the public’ can remain solvent. That is FUNDAMENTALLY the problem with our CENTRALIZED SYSTEM on the corner of Wall Street and Broad.
Here is an example for Yellen to swallow………..here is how hedge fund cheats the public using fiat instruments that are centralized and need regulation. Decentralized finance has no need for any of this. It eliminates all middlemen because it removes people from controlling the market.
Here’s the bailout play coming.
Robinhood “raised” $1B overnight from existing investors and it is getting larger.
The elites in suits and hedge funds can’t let Robinhood die this way because it harms their centralized paradigm. Who’s gonna’ help them front-run so many average Joes otherwise?
Remember the ultimate Juris maxima:If something’s free, you’re the product. Robinhood was free so the retail investors from Wall St bets are likely the product being sold to someone else. That someone else is who Biden and Yellen will protect. You can bet your ass on it. Those are the cheaters…………..and that is why we all need to unbank ourselves.
Bitcoiner’s will forget what you said, bitcoiner’s will forget how you did it, but they will never forget a centralized mistake.
Congress saved the hedge funds, but the US taxpayer can’t even get their stimulus checks. Wall Street bail outs are printed out of fiat dollars, not Bitcoin. The call for the BTC Standard is getting louder by the day.
I suppose since ‘We The People’ have seen over $20 trillion in debt monetized by Central Banks in the past decade, we’re now a bit jaded, but what the SNB (and others) is doing is MONETIZING equities. The retail investors of Wall Street Bets got fed up did their homework and put the elite Hedge fund managers balls in a vice grip to facilitate change. They are interested in destroying the centralized world of Wall Street. What did the retail investors of Wall Street Bets really do?
When investors come into a decentralized market they add mass to it. That mass creates its own value that Wall Street and the government cannot control. That is the point of the silent revolution happening now.
This is who regulators hurt most
One country’s failure to realize the opportunity with crypto is another’s gain. Deregulating Bitcoin only hurts legitimate businesses that can benefit from its global, deflationary properties. Countries that have outlawed Bitcoin forced it onto the Black Market and lost out on benefiting it themselves.
Think about marijuana and why U.S. states are now legalizing it for major tax income.
Let me reiterate, legitimate U.S. businesses are the ones the U.S. hurts most with regulation. What’s stopping another global powerhouse like China or Japan from welcoming Bitcoin with open arms?
Don’t forget that Bitcoin is a deflationary currency, something that the U.S. needs desperately right now after printing 20% of all U.S. dollars ever created. Yes, one-fifth of ALL U.S. dollars were created in 2020 alone to fuel stimulus packages and financial bailouts linked to COVID-19.
Inflation is predicted to get much worse under Biden. This is why I wrote BTC #6 for my patrons and members.
The Biggest Mistake Many Crypto Investors Make
They invest in Bitcoin without first understanding it.
They just want to make money ASAP and go home with their loot.
Sure, you can probably pull this off if this is you. The problem is, however, that when Bitcoin drops you may not have the stomach to wait it out. Your lack of confidence — of knowledge — will make you pull the trigger on what could be the best investment of your life.
Learn as much as you can about crypto before investing any money into it. This game isn’t for the faint of heart. You have to believe in what you’re getting into, or else you might burn a hole in your pocket.
WHAT JUST HAPPENED THIS WEEK IN WALL STREET IS THE MOST BULLISH THING FOR BITCOIN. REALIZE THIS NOW, folks.
2020 was when the criminals in the Deep State made their moves. 2021 is when the people counter those moves by forcing decentralized blockchains down the Deep State’s throat.
They can only control us because we use their currency. We can opt out using the people’s currency = BTC.
The early Bitcoiners you call lucky actually had 3 things:
1. Curiosity: That’s why they found BTC before you.
2. Intelligence: That’s why they grasped BTC before you.
3. GUTS: That’s why they bought BTC before you.
If you still don’t own BTC, you lack at least one of those attributes.
Computer codes are algorithms that are shaping the way we live, and not in a good way.
Google is a company who uses algorithms to collect data to take advantage of it in some way to generate profits.
An algorithm is a set of guidelines that describe how to perform a task.
So why did they buy FitBit?
They needed data to fill the data sets to run their algorithms before the enter the healthcare sector of the economy to cheat the public using code.
Medical algorithms are all code based and are being used to set up “evidence based guidelines” Why is this a problem?
Can an algorithms contain evidence based guidelines if the evidence used to build the data base is flawed toward a centralized controller who does not have health as their only outcome?
The implications might be worse than that ^^^^^.
Even within computer science, machine learning is notably ‘opaque’. Do you want something opaque ruining your treatment? How can the doctor understand something he cannot read or see? Healthcare algorithms are now increasingly central to the ways that we treat, making it all the more important to disperse that fog. Part of the issue, though, is that machine-learning algorithms are effectively programming themselves, meaning that they can sometimes be unpredictable, or even slightly alien. THE DOCTOR HAS NO CONTROL OVER HOW THEY EVOLVE. That means you are being treated by something with no medical training. Moreover, the operations algorithms contain are sometimes obscure even to those who originally created them! This means a physician needs to understand the problems with coding if they are able to stop a bad effect. Do you think physicians were trained for this?
No, they were not.
GOOGLE and Apple are part of the industrial healthcare complex and they have an “algorithm playbook” for medicine to eliminate physicians and nurses from the equations to gain control of the decision making tree to save a corporation money during the decision process……..but these computer based guidelines are not the one’s doctors would use to make decisions for your care. They are done because they drive profit and lower costs.
They help many companies in the Big tech industry to help create illnesses using technology and then they create algorithms based solutions for these said sickness and they win profits for the share holders. This is why they are the highest valued companies in the world today. They use centralized control of data to get the outcome that suits them best. It is very much how a slot machine works for a casino.
We got manipulated, my fellow doctors by “a technocratic oath” we were all sold in medical school and residency. The government helped sell this drama to us. We screwed ourselves first, and as patients got screwed as collateral damage. From Google to Apple to 23andMe, many major tech companies are getting into the health research space over the last 20 years while doctors are being replaced by midlevel providers without MD degrees. GUESS WHY?
You do not need a physician when you input data science into centralized medical algorithms. In fact this is how you transfer money from physicians to Big Tech via the centralized arm of the industrial healthcare complex. The government and Big tech want you replaced by their machine learning. Wise physicians must learn how to decentralize their practices at once.
Where are the physicians today that understand this linkage??? They are all employed by the industrial-healthcare complex. They can be found pushing the narrative of their bosses on social media helping Big Tech perform this task to hundreds of thousands of doctors who have no idea their profession is being stolen from them by a centralized system. Physicians have always had a patient’s best interest in mind and this kept them free of centralized ideas in the hospital or healthcare. Therefore, it should make sense decentralized gatekeepers should provide the research as a care ancillary at the point of care. Decentralized treatment options offer the freedoms to implement thoughtful control to benefit our patients well being and improve health outcomes. Do you want to see a picture that makes the point clear?
The chicken on the left mimics patients and physicians who are forced into the hospital cages and the care programs the industrial healthcare complex offers today.
THE TECHNOCRATIC OATH IS A KILLER FOR DOCS AND PATIENTS: OPT OUT
Is a system of centralized control mechanisms run by algorithm data grabs by the technocratic support staff (google). The use of medical algorithms was not an attempt to solve complex healthcare problems at its core. Hubris and funding without contextual knowledge are a dangerous combination. Why do physiicans keep waiting to get invited to this party when we should be the ones hosting it and driving the discussion, for our patient’s benefit? It is time for doctors to opt out and become fully decentralized if patients are to be helped escape chronic disease epidemics.
Big tech has done a lousy job of solving healthcare problems because they miss the fundamental point you just made about the doctor-patient relationship.
Today, health guidelines are built by centralized Big tech companies using algorithm as their prescription pad.
Moreover, you do not know what the code in them says or how it is programmed to harm health, you’ll never understand how your healthcare is as manipulated the stock market moves. It becomes a casino game where hospitals and centralized supply chains control how care is delivered. This is why decentralized finance programs are being built on decentralized blockchains in finance to cut down the influence and costs in the financial world. We need to stop allowing the industrial-healthcare complex to control how health care is delivered because they use centralized algorithms which cheat the public and doctors using code in those algorithms to do the job in stealth fashion. These centralized algorithms then take the ‘care’ out of health’care’.
Centralized algorithms act as gate keepers in a network and this drives inefficiency in time and value. This is decentralized blockchain systems were innovated. If there are gate keepers, they will attempt to control free markets for their own manipulation. This is why health guidelines and screening programs fail the public’s health.
As a result, today’s health guidelines on treatment and screening are mine fields. They were designed to ‘allegedly’ keep us healthy, when in reality, the companies who controlled the code, used it to enrich the profit and loss sheets of the industrial-healthcare complex. This environment resists change and keeps vested interests entrenched while the public’s health outcomes show up in a myriad of modern epidemics with no cures.
The industrial-healthcare complex is the entertainment division for the owners of centralized medical algorithms. The initial programming begins in medical school and continues on your entire career and it tries to hide itself from your everyday life. This is why cheating with code is so successful.
If you want to know why Big tech is trying to support the use of physician assistant (PA’s) and nurse practioners (NP’s = mid-level care) it is becase big tech has foster thinking in the industrial healthcare complex they can ‘harness” big data to predict the healthcare decisions through pattern identification using medical algorithms. They have not done this yet, because it cannot work.
In this new centralized system of medical algorithms, the industrial health care complex believe humans only deserve midlevel care because the code in algorithms can replace the thinking of physicians. Their logic is that most things in healthcare can be delievered by coding mechanisms according to the latest ICD code. A midlevel person is all one needs to input the data to the algorithm and disease should vanish. Under this system diseases have exploded while the industrial healthcare complex profits have exploded. All one has to do is check out their stock prices to see this effect.
They believe, that only the few chosen diseases/patient will deserve real (thinking) doctors in the new code based system.
U.S. healthcare cost is $3.5 trillion centralized, and they want to keep the gravy train running by using centralized algorithms to generate profit, not healthcare results for you.
Science is too delicate for technocratic forces to govern. It turns clinicians and scientists into salesman for the industrial healthcare complex.
The secrets of this earth are not for all people to see, but only for those who will seek them. That is the real job of a physician who is not biased by his master’s ideology.
The Robinhood’s app, today, is boosting random ‘redditors’ ability to transform the energy in money at the expense of hedge fund alpha right now. Hedge funds time to react has collapsed faster than they could adapt.
This means the momentum of monetary energy is capable of creating ripples in the space/time existence of hedge fund managers. Might this be another powerful example of how DeFi will force changes in the reality we all experience? Is this another reason for hedge funds might have to use BTC as a Treasury hedge to gain more time back to make more money = alpha while compressing time?
This issues with Redittors, GME/AMC, hedge funds, and Robinhood show that Web 1.0 and 2.0 are centralized. We need a decentralized web. If we had this Parler could not have been shut down. Today, Robinhood shut down to lock out the Reddittors.
The public has an experienced history with the barons of Wall Street. Time is never what it seems to be with them. The Street never seems to lose in this casino game. Is regular Joe is getting more pissed as time elapses and ready for the revolution of money to begin?
So what happened? Citadel is an investor in Melvin Capital, which got run over by Wall Street Bets on the sub Reddit. Citadel Hedge fund is also Robinhood’s biggest customer. Robinhood was never what it pretended to be. It built its business on selling Wall Street Bets order flow to the hedge fund Citadel. In the moment of truth, we learned which customer it cares about the most. It was not the little guy.
Robinhood shut down Wall Street Bets to benefit their customer and destroy a market play.
Wall Street Bets did to the hedge fund suits what the suits have been doing to Main street Joe’s for a century. Then one call to Reddit, one call to Discord, one call to Robinhood… It there anyone out there who still doesn’t think the system is rigged against the little guys?
When the Dow Jones falls below 5%, the market stops trading. Conversely there is no 5% cut off when the market gains. That’s not a free market. That’s an SEC safety net (aka socialism) for the rich.
Should we assuming that the next time a hedge fund starts to make too much money shorting and destroying a business, that they will be de-platformed from their Blomberg terminal and throttled by their prime broker in the name of orderly markets and consumer protection? That will never happen until all DeFi applications are DECENTRALIZED!
The TIME HAS COME and this is the perfect storm for change.
Ask yourself, is true reality timeless?
For 500 years “time” has been paradoxical for physicists.
The distinction between the past, present and the future is an illusion, this comes from very well-understood physics, in Einstein’s case general theory of relativity. Is this true? Is it wise to believe time is a fixed constant?
Wells Fargo executives robbed their own customers in their mortgage scams, but let’s continue talking about some redditors shorting actions on GME/AMC. Shorting is how one collapses time in the stock market. When a price rises quickly time vanishes and value is stolen. It appears, from this perspective, reality is really a fucking illusion.
If you’re feeling rattled about learning that the stock market is a scam from the GME/AMC situation, wait until I tell you that money and time are illusions to, according to the laws of physics.
The GME and AMC show you fiat can only exist if time and value are illusions.
Is this this true?
Value is an illusion buried in the trust that makes a currency. What about time?
The paradigm says, time is but an illusion created by beings with limited perception of what reality is all about. Few people realize time is a concept that defines the past and future. Time has no place in the present, hence it might be an illusion of the human brain to make sense of the chaos we sense.
Wall Street has forgotten their own axioms when it comes to time. We should never make one customer our total priority when all you are to them is an option. With respect to time, I think we get what we deserve.
When the Fed prints money and steals time and money from you, remember: Currency fluctuations, inflation, and paper hike. You may not control all the events that happen to you by the Fed, but you can decide not to be reduced by them. This is where BTC enters the equation. We all need to choose wisely how to limit the effect of fiat on our lives. Will this decision affect time in our life? Understand how BTC controls the flow of value and time in your world. Be brave now: and issue a letter to all your clients that you are hiking the prices. Most clients will respect you for it, and those who don’t were never going to value you appropriately either. They are just like the Fed.
Pricing is actually a pretty simple and straight forward thing. Customers will not pay literally a penny more than the true value of the product you serve. You know you’re priced right when your customers complain—but buy anyway.
What I ‘charge’ today has nothing to do with yesterday or tomorrow. It has to do with ‘now’!
The present moment is where time is timeless.
Price is what you pay. The value of time is what you get in the transaction. I just completed my first crypto brainstorming session this week with a small group of people who wanted to to pick my brain to sample my value to expand the space time in their life they experience. That made time and value tangible to them.
Today, forget the prices of things and cherish the value of people.
Others value your time exactly as you do. If you value it, so will other people.
If you don’t have a plan for your week before it begins, you’ve planned to fail.
Leaders who value their time always start with a plan for the week before it begins.
Always have a plan for your week before other people plan it for you.
Spend 80% of your time with people who give you 80% of your results and stand back and be amazed at what happens. I use the filter of social media to figure out who that 20% is.
Regardless, if you haven’t decided ahead of time the kinds of people you’ll brainstorm with, you’re sunk.
The laws of nature are timeless, but the concept of time and value are not. If the physical laws of Nature can change in time, then there’s nothing beyond time.
So what do I have to say about time now? Time is real. Based on the best evidence we have, and everything which appears real and everything which is true is real or true in a moment, which is one of a succession of moments. That’s what we experience. And the question should be in this case: Is that the structure of nature? Does nature exist in a series of moments, one after the other? Is that what’s really real about the world? Or is that, as Einstein said, an illusion, and there is some timeless picture which is the truer picture?
I think the present moment is built for BTC. I think fiat is in our past. Why does time confabulate with our ability to reason? I personally believe we are missing a historical dimension to physics and this is why time confounds us. As such, I believe, perhaps laws of nature change as the environment change.
BACK TO REALITY TODAY:
I think this is why what has happened to GME and AMC has happened. I think the General Motors recent bankruptcy was the wormhole that opened this can or worms. GM’s bankruptcy seemed to suspend time because of how it changed the rules of how bankruptcy were supposed to work. When GM’s death was put off it was as if time was suspended, and in th bankruptcy procedings, equity stakes were kept alive for the sake of “time”…………and the reason why TIME was dilated in this way followed no laws; it was done for the survival of pensions for those whose employment for GM had long expired. The laws were changed to preserve the TIME and value of past workers. The lesson? When value collapses in pensions, so does time. This is why it is now time for a new version of people’s money.
The quick take home: You’re only self sovereign if you run your full node on a miniaturized backyard molten liquid fluoride salt thorium nuclear reactor. Sounds crazy? Read on.
When you read the White paper of Satoshi and the original ETH papers the one bad thing about both coins is the energy required to mine coins. This process will become harder and harder as we approach 2140. As we get closer to 2140 the process will require more energy. In this way, you should look at BTC mining like trying to climb up Mount Everest. When you are young it is easy to get to base camp. It is tougher to get to the Summit even when you are young. When you are older just getting to base camp seem ridiculous. When you are above 50 years old getting to the summit is nearly an impossible task because you do not have the mitochondrial power to get the job done with a falling oxygen tension. Mining BTC from 2050-2140 will be like trying to climb Everest at 60 years old. Analogy over.
ETH 2.0 is trying to solve this issue by changing from a Proof of Work (PoW) blockchain to a Proof of Stake blockchain (PoS). Proof of stake (PoS) differs in that instead of miners, transaction validators stake crypto assets they own now for the right to verify a transaction. They make money in fees to process those transactions. These validators are selected to propose a block based on how much crypto they hold, and how long they’ve held it for. this means ETH 2.0 favors miners with a lot of skin in the game of ETHER.
Other validators can then attest that they have seen a block. When there are enough attestations, a block can be added to the blockchain. Validators then are rewarded with cryptocoins for the successful block proposition. This process is known as “forging” or “minting”.
The main advantage of PoS is that it is far more energy-efficient than PoW, as it decouples energy-intensive computer processing from the consensus algorithm. It also means that you don’t need a lot of electricity for computing power to secure the blockchain.
One of the reasons BTC blockchain is not being used for transactions as much as it could be is tied to the speed in which it works. Speed is sacrificed for security. The security of the BTC network is second to none. This means financial institutions may favor one over the other initially. Right now institutions are favoring BTC. The “Ethereans” are betting large this will change when ETH 2.0 programming is shown to be successful on delivering its promises.
PoW = Proof of work
PoS = Proof of Stake
In ETH 2.0 this is exacerbated by the fact that only one miner will find the correct hash, so the work done by all the others is not actually used. As a result, PoW consensus requires a huge amount of computational power and electricity and is very inefficient. PoW early on in a block chain life is like being elderly and trying to climb a small hill. Tough but doable. The programmers building ETH 2.0 realized this limitation with PoW blockchains and said it should be easier for miners now if we want a faster adoption to build out the Monopoly board on top ETH 2.0. For this sole reason, PoW was changed for PoS. PoW protocols are important because they enabled the creation of early blockchain networks, but they are very inefficient from an energy standpoint too. In the current political environment this will also slow adoption unless it is improved.
CRYPTO ENERGY COSTS
In 2019 each transaction on the Ethereum network uses about 29 kwh (Kilowatt hours), enough electricity to power an average American household for 24 hours! The electricity used by Ethereum mines could power 652,669 American households every year. Ethereum mining annually uses about the same amount of energy as the country of Bolivia.
Is there a way to mitigate this energy risk today they we are ignoring?
Yes. BTC miners are dominated by China because the western provinces of China have given miners cheap sources of electricity due to overbuilt hydroelectric power plants.
What should new countries who mine due to capture mining capability from these people?
They should switch to a clean nuclear power but not today’s version of nuclear power that came from WW2. They need to use thorium as their fuel source.
Why do I suggest this?
The market will determine this………..thorium is the cheapest way to create electricity on Earth. In fact, it is the way nature makes geothermal power of vents, Earth quakes, and volcano’s.
Thorium is the element in the Earth’s crust that drive geothermal vents everywhere on Earth. It is unending source of power that drive plate tectonics. Without massive amounts of thorium in our crust, our planet likely would wind up like Mars. A dead red dessert with no volcanos and no plate movement. Without thorium the interior of your planet cools too fast and without molten lava you have a lowered magnetic shield to protect you from the sun. As the sun’s power blasts a planet its cathode rays strip a planet of its atmosphere. See the Moon and Mars as exhibit one and two.
Thorium is very common element in the Earth crust at 10 parts per million. We have an unending source of it everywhere in the world. It is cheap to mine and its half life is 14 billion years. This is same length of time we believe the universe has been around. This makes it hardly radioactive at all.
U.S. equity markets continue to be the most favorable listing venue for mining companies. US miners have a big advantage over Chinese miners because they can raise through [at-the-market] offerings, which provide a very solid financing method for public companies looking to scale up their operations. Foreign companies have more limited financing levers and have a relatively harder time raising capital through equity.
A prominent example is Nasdaq-listed Marathon Patent Group (MARA), which recently spent $50 million on a fleet of Bitmain’s state-of-the-art S19 Pro Bitcoin mining computers earlier this year. Marathon is building out a 105-megawatt (MW) mining facility in Hardin, Montana, as part of a venture with Maryland-based power provider Beowulf Energy. Beowulf Energy is a private company linked to new ways to produce energy.
What don’t most people in the crypto-world know? A lot of the power production in the U.S. has been deregulated, and private equity or hedge funds own a lot of power-generation facilities. These hedge funds are now trying to set up colonial positions in BTC and ETH. They want to make money on coin production as they add crypto assets to their clients portfolios. Some of the best ways to generate alpha is to produce your own coins at the lowest cost. That is how billions can be made. So, the wise, follow the money to see where the market is headed. There is a lot of signals that thorium will be used to make the USA miners the strongest positioned groups until 2140.
Guess why MARA picked Montana to build this brand new BTC mining region?
Do you know who Glenn Seaborg is? If not, he is the reason MARA has chosen Montana to mine.
Glenn Seaborg was one of our great scientist who worked in the metallurgic labs during the Manhattan Project. He won a Nobel Prize. He figured out how to make plutonium quickly from uranium to build nucelar weapons. He discovered plutonium in this quest. He also stumbled into a solution for energy production for man using thorium based nuclear fuels. The problem was the military focus was on building bombs instead of building a cheap source of energy for the planet.
The story of the thorium-fueled molten-salt reactor effort at Oak Ridge National Laboratory, strongly motivated by the discovery of a series of books (Seaborg and Loeb (1981), Seaborg and Loeb (1987), Seaborg and Loeb (1993)) written by Glenn Seaborg and Benjamin Loeb covering Seaborg’s years as the chairman of the US Atomic Energy Committee in the 1950s.
In the last of these books, The Atomic Energy Commission Under Nixon (Seaborg and Loeb (1993)), Seaborg describes budget cuts early in the Nixon Administration (1968-1971) that led the AEC to curtail research into breeder reactors. This is the same time the dollar was moved off the gold standard to boost the economy using money printing. How is that for irony!
Seaborg described in the books how Alvin Weinberg, head of the Oak Ridge National Laboratory (ORNL) and ardent proponent of the thorium- fueled molten-salt breeder reactor (MSBR), argued for the continuation of that line of research because of how cheap energy could be made safely.
The problem was that a new type of reactor needed to be built at a time where Nixon removed the US dollar from the gold standard because of a slowing economy. This sunk the manufacturing effort. Seaborg goes on to describe how the MSBR program was cancelled in favor of the plutonium-fueled, liquid-metal fast-breeder reactor (LMFBR) but concedes later that that may have been a mistake. (Seaborg and Loeb, 1993, pg.179).
Seaborg saying this in a book is a mouthful because he won the Nobel Prize for discovering plutonium and the high pressure nuclear reactors that are used today in the US for electricity production.
The cheapest source of electricity on Earth is to mimic what thorium does in the Earth’s crust. Doing so allows us to manufacture an unending source of energy from the thorium atom, at a fraction of today’s cost. This is what drives geothermal cycles on Earth. See Iceland and Hawaii as example 1 and 2 for that awesome power display. The tectonic plates have moved on Earth for 4.6 billion years while Mars and the Moon have long ago stopped.
Why is this a big deal? I got into a heated debate with one of my Farm members in the energy business in 2020 and he told me he bailed on trying to mine BTC because of the energy costs. He also told me that he was not interested in BTC deeply because China dominated the mining scene. I told him I thought he was being myopic. This blog is being written as my response to that idea.
China might dominate crypto mining presently, but that won’t be true much longer. China may have lorded it over the crypto mining space until now thanks to cheap labor and a massive over-build in dam and hydro generation infrastructure. But the U.S. has begun catching the attention of Chinese players looking to diversify. In fact, Chinese miners have called on the CEO or Argo Blockchain to help them set up a transfer of mining stake to the USA. Why is that?
Miners are always looking for more political stability, which North America offers, and power and hosting costs in North America are competitive and sometimes even cheaper than Chinese options. How could they be cheaper? Private energy companies are now going to use new thorium technology developed by Glenn Seaborg during the Manhattan project to generate power that is 90% cheaper than China’s CCP can deliver.
WOW………..How?
NUCLEAR PHYSICS LESSON:
Seaborg recognized that for each absorption of a neutron in uranium-233, it would produce more than 2 neutrons. This was the key to its ability to operate as a breeder reactor. Each neutron produced could split another U-233 atom to unleash energy in a nuclear fission reaction. It turns out any nuclear reaction can be sustained as long as the process liberates at least two neutrons. The best chance to make an unending source of energy is a reaction capable of generating anything above 2 neutrons. Seaborg knew that the key was which elements on the period table produced the most neutrons when they themselves were hit with a neutron (fission) in a reactor. But how much more than two was the question? the higher the number of neutrons released = the more energy can be harvested from the fission of an atom.
When Thorium was tested it was found to be ideal to be a neutron breeder for the nuclear reaction because its fission elaborated 2.35 neutrons per split. With its 14 billion year half life, it became the best candidate to make electricity cheaply. The problem was the military was not interested in electric power, they were interested in explosive power. To make massive amounts of electricity from thorium, thorium needs a special reactor to do it. Earth seems to do it naturally by some mechanism in the core of the planet. This reaction drives plate tectonics for 4.6 billion years.
This means that thorium can be an unending source of energy to generate electricity and it will be quite cheap to do so with minimal radioactive waste. This idea was born during the Manhattan project but it was abandoned for the fission use of plutonium instead. Genreral Groves made this decision because the military at the time was only interested in making a bomb, and not a super power source.
The largest thorium vein deposits in the world are in the USA. The largest volume known high-grade thorium resources are in the United States. China does not have much thorium in their landmass. As a result, they must purchase it.
Two thorium vein districts-the Lemhi Pass district of Montana-Idaho and the Wet Mountains area of Colorado dominate the known high-grade thorium reserves in the United States.
Now you can see why USA cryptominers are going to Montana to set up shop.
The obvious geopolitical implication of this move is that the U.S. could eventually take on China in this nascent arena. The cryptomining community would rather couch this in terms of greater decentralization, whether that means geographical spread or selling mining company shares to the public. I see things differently.
Before the US sets the Bitcoin standard for its Treasury it must guarantee the network of miners can not fail to ensure the 51% rule is never violated. Therefore, the goal is not for the U.S. to dominate bitcoin mining. If you understand the Satoshi white paper that is not the point, and it’s not gonna happen. The goal here is to decentralize the blockchain throughout the world to make sure no one country to get to the 51% limit.
That is why American miners have begun to think way outside the box. None of them have admitted why they are going to Montana…….but I figured it out because of my deep understanding of American history.
Interest in thorium was reborn in the military in the 1950s when the Air Force wanted to build a nuclear powered airplane. As crazy as this sounds today, the molten salt reactor using thorium would have never been built if the military did not ask Seaborg and his crew at Oakridge labs to see if this was feasible. This project done fifty years ago, created the era of operating molten-salt reactors came to a fast conclusion with the shut down of the Molten-Salt Reactor Experiment in the 1950s. This was the only time thorium nuclear reactor was ever built by the US military.
Today, China is trying to build fifty of them using stolen US IP from the Manhattan Project. China has a big problem, however. Their crust is relatively thorium poor, not because China does not have it. It is because it is present in hard to reach parts of China or heavily populated regions. Thorium deposits have been recognized in all the first level tectonic units in China but are mostly concentrated in the northern margin of the North China Craton, the Central Asian Orogenic Belt, South China and the Mian’ning-Dechang metallogenic belt.
HOW IS ENERGY UNLEASHED CHEAPLY FROM THORIUM? NEUTRONS ARE THE KEY
When we use a neutron to hit a nucleon of thorium it releases 2.35 neutrons in core. We learned this right around FDRs death in April of 1945. At this time, only the United States would possess the knowledge that uranium-233 could, for all practical purposes, ”catalyze” energy release from a supply of thorium indefinitely.
Read that again folks……….
Thorium can create energy longer and more efficiently than any other source of energy on Earth. Seaborg is the guy who figured this out during the Manhattan Project.
This implies it might be the ideal answer to fossil fuels and the climate change debate as well.
The person who discovered plutonium (Seaborg) realized this in 1942 when his graduate student did the experiments and the mathematics to prove it. He famously told that grad student that this energy discovery would be worth 40 quadrillion dollars in 1942. He wasn’t joking. Nuclear fission is 1 million times as energy dense as a chemical reaction in releasing energy.
******Because of this history lesson, I will make a large prediction. The adoption of crypto assets will change the climate debate and how we view nuclear power plants. I believe BTC and ETH mining energy use will make us (THE USA) realize we need to use thorium breeder fuel with molten liquid Fluoride salts for reactors and the entire power grid of the world will change before 2140.
I think this will be a great unintended consequence of crypto adoption. This will change the world because it will increase the thermodynamic efficiency of power generation and this will effectively end the use of fossil fuels. It will also allow miners from 2075-2140 efficiently get those last coins out that are likely going to be worth trillions of dollars.
The C-H bond in fossil fuels has a million times less energy in it than the Thorium nucleus does for fission.
This is why the Earth’s core remains energy vibrant given its planetary mass, while Mars furnace has turned dormant. It’s crust is thorium poor, so it cooled faster than the Earth and this is why it has no magnetic field production. Thorium powers the molten core of the Earth too to create our magnetic field!
On Mars, the region of highest thorium content, shown in red, is found in the northern part of Acidalia Planitia (50 degrees latitude, -30 degrees longitude). Areas of low thorium content, shown in blue, are spread widely across the planet with significant low abundances located to the north of Olympus Mons. Olympic Mons is a massive super volcano, the largest in the solar system that burned out billions of years ago. The fact that it is present where thorium is now low is no surprise considering Mars is a dead red desert.
MANHATTAN PROJECT LINKS
Seaborg was a Nobel Laureate and one of the founding fathers of the atomic age. He was co-discoverer of plutonium and later served as chairman of the Atomic Energy Commission. He worked with a scientist named Wigner who was the one person in history, who first realized the power of thorium to generate energy.
Seaborg’s interest in fluorination as a separation technique came from the fact that thorium, when exposed to neutrons in a reactor, would absorb some of those neutrons and form small amounts of uranium-233. If the thorium and uranium were chemically converted to tetrafluorides, which is a rather simple chemical step, and if that mixture was further fluorinated to convert the uranium to a hexafluoride, then it would be rather easy to separate uranium from thorium.
In 1946, the public first became informed of uranium-233 bred from thorium as “a third available source of nuclear energy and atom bombs” (in addition to uranium-235 and plutonium-239), following a United Nations report and a speech by Glenn T. Seaborg. Few people have asked why the thorium cycle was abandoned.
U-235 is the world’s primary nuclear fuel and is usually used in high pressure light water reactors. Seaborg was the scientist who developed these nucelar reactors and holds all the patents on them. The high pressure use is what usually leads to nuclear accidents.
This path was chosen by the military because it could be used to make plutonium fastest to create a nuclear arsenal while also generating energy. Today, that decision seems foolish when you understand that thorium reactions produce an unending source of heat to create electric power with a much lower toxicity profile.
Th-232/U-233 is best suited to molten salt reactors (MSR). These reactors have produced electricity and nuclear bomb material but have the highest risk level of core meltdown because they all use higher pressure systems to operate. Thorium can operate in lower pressure systems to liberate massive unending sources of energy from the thorium nucleus.
Their radioactive decay produces about half of the Earth’s internal heat. That heat, in the form of geothermal activity comes from the action of thorium on uranium in natural breeder reactions in the mantle and core.
Thorium can be used as a fuel source in liquid salts of fluoride. It appears this is how the dynamo of the Earth appears to work in Nature. The liquid fluoride thorium reactor (LFTR; often pronounced lifter) is a type of molten salt reactor. LFTRs use the thorium fuel cycle with a fluoride-based, molten, liquid salt for fuel. My bet is that inside the Earth their is more options to use than fluoride.
Reactors that use the uranium-plutonium fuel cycle require high pressure fast reactors to sustain breeding, because only with fast moving neutrons does the fission process provide more than 2 neutrons per fission. With thorium, it is possible to breed using a thermal reactor. These reactors produce more thermal energy and that is why they can produce electric power very cheaply when thorium is in deep supply.
This was proven to work in the Shippingport Atomic Power Station, whose final fuel load bred slightly more fissile from thorium than it consumed, despite being a fairly standard light water reactor. Thermal reactors require less of the expensive fissile fuel to start, but are more sensitive to fission products left in the core. This is the massive pay off that American BTC miners are looking to exploit in Montana.
Despite Seaborg’s interest in thorium and uranium-233, Seaborg’s overwhelming priority was the chemical separation of plutonium from uranium and fission products in the production reactors that would be built in Hanford for the Manhattan Project. This was done because the military was interested in weapons grade material only at the time. They did not care about thermal power of electric power generation.
WEAPONS GRADE PLUTONIUM WAS THE GOAL NOT ELECTRICITY
Fluorination was not a very effective technique for separating uranium from plutonium, because plutonium had a variety of valence states and could also form a gaseous hexafluoride. If plutonium hexafluoride formed in preference to uranium hexafluoride then there might have been interest, but just the opposite took place. During fluorination all of the uranium would react with the fluorine (forming gaseous UF6) before the plutonium would, meaning that an entire volume of uranium parent material would need to be fluorinated to UF6 before the plutonium would begin to fluorinate to PuF6. This was undesirable, since they estimated that the Hanford reactors would have a million grams of uranium in their exposed fuel for every thirty grams of plutonium.
The fact that uranium would fluorinate to an easily-separable gas and thorium wouldn’t, made fluorination an attractive chemical separation technique for a thorium- uranium-233 reactor, but not attractive for a uranium-plutonium reactor.
Since the priority of the Metallurgical Laboratory was chemical separation of plutonium, fluorination assumed a role of lesser importance over time. thorium was a better fuel to generate electric power and Seaborg transmitted this idea to Weinberg as his student in the 1940s.
Weinberg later became head of Oak Ridge Nation Labs where the concept was tested in an experiment.The LFTR concept was first investigated at the Oak Ridge National LaboratoryMolten-Salt Reactor Experiment in the 1960s, though the MSRE at this time did not use thorium as a fuel source. The LFTR has recently been the subject of a renewed interest worldwide.
At Oak Ridge National Laboratory in the 1960s, the Molten-Salt Reactor Experiment used U-233 as the fissile fuel in an experiment to demonstrate a part of the Molten Salt Breeder Reactor that was designed to operate on the thorium fuel cycle. Molten salt reactor (MSR) experiments assessed thorium’s feasibility, using thorium(IV) fluoride dissolved in a molten salt fluid that eliminated the need to fabricate fuel elements. The MSR program was defunded in 1976 after its patron Alvin Weinberg was fired by the Carter Administration in its first year. Weinberg was a student of Seaborg and Wigner after the Manhattan Project. He learned everything these two found while building the bomb.
In the last few years, the long-term strategy of the nuclear power program of India, which has substantial thorium reserves, is to move to a nuclear program breeding uranium-233 from thorium feedstock. This would be another ideal place to mine BTC/ETH for cheap power.
In 1993, Carlo Rubbia proposed the concept of an energy amplifier or “accelerator driven system” (ADS), which he saw as a novel and safe way to produce nuclear energy that exploited existing accelerator technologies. Rubbia’s proposal offered the potential to incinerate high-activity nuclear waste and produce energy from natural thorium and depleted uranium. Recall, that this was under Clinton and his wife was recently involved in selling a lot of uranium stores of the USA to Russia in the Obama administration under the guise of secrecy as Secretary of State.
Kirk Sorensen, former NASA scientist and Chief Technologist at Flibe Energy who went to the same engineering school as my son, has been a long-time promoter of thorium fuel cycle and particularly liquid fluoride thorium reactors (LFTRs). He first researched thorium reactors while working at NASA, while evaluating power plant designs suitable for lunar colonies. This is big deal for space travel. The reason the space program needs to make a pit stop at the moon is to fuel themselves with thorium at the moon to get to other planets in the solar system. This is something NASA, SpaceX, and the CCP do not want people to know.
Thorium is concentrated mostly in one area of the lunar nearside, in the around the great big Imbrium basin. All space programs want to set a lunar base there. Now you know why.
In 2006 Sorensen started “energyfromthorium.com” to promote and make information available about this technology. It is believed that China has gotten most of the IP on thorium cycles directly from his website to design their own molten core reactors to be made for the Moon and for electric power production.
Nuclear fission produces radioactive fission products which can have half-lives from days to greater than 200,000 years. According to toxicity studies, the thorium cycle can fully recycle actinide wastes and only emit fission product wastes, and after a few hundred years, therefore, the waste from a thorium reactor is less toxic than the uranium ore that would have been used to produce low enriched uranium fuel for a light water reactor of the same power. Other studies assume some actinide losses and find that actinide wastes dominate thorium cycle waste radioactivity at some future periods.
Thorium-cycle fuels produce hard gamma emissions, which damage electronics, limiting their use in bombs. This is the reason the military did not use thorium in the Manhattan Project.
Thorium is estimated to be about ten times more abundant than uranium in Earth’s crust. Using molten liquid breeder reactors, known thorium resources can both generate world-scale energy for thousands of years at very low cost in a safe manner.
A 2011 MIT study concluded that although there is little in the way of barriers to a thorium fuel cycle, with current or near term light-water reactor designs there is also little incentive for any significant market penetration to occur.
That incentive likely will be BTC mining if coins are valued in the hundreds of millions or trillions of dollars in 2050-2140.
They were blind to the crypto angle when the paper was written. As such they concluded there is little chance of thorium cycles replacing conventional uranium cycles in the current nuclear power market, despite the potential benefits right now in 2020. I have an idea this might change because of Bitcoin.
SUMMARY
Molten salt nuclear reactors using liquid thorium fluoride salts is the future for BTC miners. Any country that wants to get into mining needs to look into this technology. It will create the cheapest source of electricity on the planet. You can burn thorium forever with fluoride and liberate energy from a massive nucleus the way the Earth does with little risk. This maximizes E=mc^2 for BTC.
North America is now on a precipice of real growth thanks to its regulatory certainty and the huge amount of infrastructure built in the 1970s and 1980s in anticipation of growing manufacturing that never came. Now that people are starting to realize crypto mining is not some shady enterprise, the U.S. is better positioned at the boardroom level than any other country in the world to take advantage of the trend.
The operational costs are a little bit more expensive in the U.S., but when you’re sinking $100 million or even a billion dollars into an ecosystem for infrastructure you’re looking at stability. China does not offer stability that the US can.
Some government support would also be helpful here and I think the Treasury moving to a BTC Standard will facilitate this change. To this end, Core Scientific has put together a policy paper and has been working with the Chamber of Digital Commerce to get the word to the U.S. government why this makes geopolitical sense and a lot of monetary sense.
We want the folks in Washington, D.C. to understand that digital asset mining is not bad and that there’s a right way to do it. Many into crypto mining are ‘greenies’ at heart. I think the right way is through renewable power sources done at a global scale. The larger that becomes, the lower the burden on the environment and it can be done to re-power the US economy and Treasury.
Osteoporosis is a disease where bones lose their strength, very similar to how bone broth breaks down bones and makes edible.
What is the home remedy to help this?
Home Remedy notes for you.
In this note I will update home remedies that are tried and true for treatment of diseases I see in clinic. I will continue to add them as requests come in. Check back in every so often. First one I get so many questions on is osetoporosis and osteopenia for patients. Here is the best up to date info of how to solve your situation without the Rx drugs that many of my collegues like to use. They are OK but for me I dont like their side effect profiles. And if you want to know more about the disease see my other note in 2010 on osteoporosis. It is more in depth about the disease and the drugs used to treat it. This note is about how to do it naturally without a prescription.
SUNLIGHT IS FREE. Get your skin in the game. Use sensible solar exposure as I laid out in the solar callus blog here on Patreon.
Vitamin D3–using a dose sufficient to raise our 25-hydroxy vitamin D blood level to 60-70 ng/nl. I’d much rather people use sunlight but for those who chose to short cut nature I advocate buying the D3 from a store at 5000 IU doses. Make sure the oil base used in pill is not an omega six. This is harmful for the skin. I base initial dosing on current lab values so it does require a blood draw. If you want your PCP to draw it and send the numbers to me to give you a consult let me know. I do not like the 50,000 IU a week dosing that the Rx vit D3 has.
Vitamin K2–Borrowing from the Japanese experience, in which vitamin K2 is used as a prescription “drug” for treatment of osteoporosis (Glakay, Eisai Pharmaceuticals), we started 1000 mcg of a mixture of K2 forms, MK-7 and MK-4. YOu can also buy vitamin K2 5 mgs from drugstore.com or from lef.org. Fermented cheeses from Europe and Natto are the best source of this vitamin naturally and neither are staples in the Standard American diet.
Magnesium–Based on evidence that magnesium strengthens the bonds in bone tissue, we started magnesium glycinate, 400 mg per day (200 mg twice a day), a form that is well-absorbed and causes little or no loose stools. I prefer eating three raw oysters per day to get your Magnesium. For bad osteoporosis I advocate adding a 400 mg dose about 90 minutes prior to sleep too. Our diet in this country is very very low in Magnesium so if your overweight too add the evening dose. And if you’re going paleo/keto/carnivore dieting as well add the magnesium to slow the development of hard stools. Magnesium citrate liquid can also be used. You’ll thank me. So will your rectum.
Progesterone–We have added natural topical progesterone cream formulated for by a compounding pharmacy, 60 mg twice a day. Yes progesterone helps bone formation and it helps women sleep alot better. You can go up to 100 mg orally too under the direction of a physician.
Strontium ranelate given orally at a dose of 2 g daily appears to reduce the risk of vertebral fractures rapidly, effectively, and safely among postmenopausal women with osteoporosis. This mineral increases our C -teleopeptide cross links to make the newly laid bone down not suseptible to remodeling and how it works remains unknown but what we do know is those who use it have faster bone formation and more bone formation and the bone laind down sticks around a long time.
Exercise should be done, but one needs to be careful with it. As a bare minimum………Weight training ten minutes three times a week. Pure weights is best. Not a machine. It builds bone better than any exercise and it should be done outside using tan through clothing. The amount you start out has to be in tune with your body status at start.
That is my home version for the initial treatment for osteopenia for men and women. We also advocate getting your hormone and vitamin D levels assessed during this treatment for metabolic bone disease. If you need more help consider becoming a member and I would be happy to help you out ——–>>>>kruseatdestin.com
The Ethereum network is one of the largest and most widely used blockchain networks in the world. in 2021, it is beginning to meet its promise of supporting all the DeFi projects that Web 3.0 promises. It is also one of the world’s largest open source projects with go-ethereum being one of the largest projects on Github. Since its first release in 2015, Ethereum has been making strides towards its ultimate goal of becoming a decentralized global computer that could one day replace many of the centralized applications that we use in everyday life.
So where do we stand right now in the crypto ecosystem?
ETH 1.0 lasted from 2015 to December 1, 2020 and included 3 stages. ETH 2.0 was released 5 weeks ago and is the fourth stage of the blockchains evolution. Ethereum 2.0 is also called Serenity in some circles. It will be the largest and most comprehensive update ever implemented by the Ethereum community. That community makes up over 3500 of the best computer programmers in the world.
ETHER’s Blockchain has had 4 stages of development.
The first stage of Ethereum was called Frontier. Released in 2015, this was the first live version of the network. Frontier allowed users and developers to mine Ether, build Dapps (DeFi apps) and tools, and experiment with the new platform. Most of the development here went to gaming. Many ideas where beta tested in this phase.
The second stage of Ethereum’s development was Homestead. This update was the full production stage of ETH 2.0. What did it do? It provided many improvements to protocols that will become the foundations of future upgrades and increased transaction speed. The BTC blockchain is a slower chain due to its design and purpose. It takes a minimum of 10 minutes to make confirmations in its blockchain. With Homestead ETHER goal was to get to 15 seconds per confirmation increasing transaction fees tremendously.
The third stage of Ethereum’s development was called Metropolis and this ended on December 1,2020. Metropolis was divided into two releases, Byzantium and Constantinople. Byzantium was released in October of 2017 and was lighter, faster and more secure than the previous versions. Some specific upgrades include more predictable gas prices (cost of transactions on ETH2.0) and easier programming for developers to build out the DeFI space on the ETH 2.0 Monopoly board. Byzantium also improves privacy. Many of the Bitcoin maximalists hammered ETHER because it did not have as tight as privacy as BTC so many people thought it would fail because of this. So far this has not been the case as of January 2021. In fact, in December 2020 ETH 2.0 made over 6 billion dollars per day of transactions safely.
The single most important issue currently facing the Ethereum network is scalability. The Ethereum network supports hundreds of decentralized applications and needs to process a high number of transactions per second. Adding more nodes does not increase the scalability of the network because each node needs to verify each transaction. Increased use of the Ethereum network, including the creation of more Dapps and the execution of more transactions, has led to an on-going increase in the time and cost of transactions. Here in lies ETH 2.0 greatest risk.
The cost of gas on ETH 2.0 since December 1, 2020 has been a hot top in the ETH community. I posted about that in the last blog. I think we need to be cognizant of this. By no means do I think this invalidates ETHER as a part of your portfolio. I think you need to be aware of the risks that ETHER has.
The Ethereum blockchain processes about three to four times as many transactions as Bitcoin. It’s still not enough, however, to meet rising user demand for the cryptocurrency and prevent network congestion. We can see transaction speed off each chain right now using this : HYPERLINK
Why is this a big deal? ETH 2.0 is aiming to replace Paypal and Visa in the DeFi space. Can they do it right now? No. Why? Ethereum network runs slower and less convenient to use right now. In a speed test run in 2021, the network only managed 15 transactions per second (various sources have stated anywhere between 12 and 45 transactions per second over the last year). In comparison, centralized networks such as PayPal and Visa regularly perform 193 and 1667 transactions respectively, and Visa’s maximum capacity is close to 24,000. For the network to achieve mass adoption, efforts to massively increase scalability are central to the development of Ethereum 2.0.
How are the programmers going to fix this? After investigating several other options, the Ethereum team settled on a process called sharding as the best solution for network scalability. Off-chain solutions like plasma chains and state channels will also likely be used to take load off of the main network and further increase scalability. No one knows if this will work as of January 2021.
What are some other risks?
Another major issue that will be addressed by ETH 2.0 is efficiency and environmental sustainability. Like Bitcoin and many other blockchain networks, Ethereum currently relies on a proof of work (PoW) consensus protocol to ensure the security of its network. PoW systems involve a number of miners who own computer hardware connected to a blockchain network. Miners are rewarded for putting their hardware to use by the network.
A major benefit of PoW systems is that verifying whether or not a new block belongs to the chain is a very fast and relatively simple process. However, in order to ensure the security of the network and prevent fraud, it is computationally expensive for miners to find the correct hash input. What are the legal risks of screwing this up?
Read this paper from crypto lawyers. It appears this is a big issue for the blockchain, the developers and the exchanges who use BTC or ETH 2.0 for transactions. This aspect o f property flow through blockchains is something to monitor. https://craigwright.net/blog/law-regulation/the-property-flaw-of-lightning/
One of the reasons BTC blockchain is not being used for transactions as much is the speed in which it works but the security of this network is second to none. This means financial institutions may favor one over the other initially.
In ETH 2.0 this is exacerbated by the fact that only one miner will find the correct hash, so the work done by all the others is not actually used. As a result, PoW consensus requires a huge amount of computational power and electricity and is very inefficient. PoW protocols are important because they enabled the creation of early blockchain networks, but they are very inefficient from an energy standpoint too. In the current political environment this will also slow adoption unless it is improved.
In 2019 each transaction on the Ethereum network uses about 29 kwh (Kilowatt hours), enough electricity to power an average American household for 24 hours! The electricity used by Ethereum mines could power 652,669 American households every year. Ethereum mining annually uses about the same amount of energy as the country of Bolivia. This is obviously a major issue that is actively being managed with ETH2.0.
This, right now is the biggest risk to ETH 2.0 in my opinion in January 2021. To solve this problem ETHER community is moving from a PoW to PoS design transition. ETH 2.0 is in design to transition the network from a proof of work protocol to a proof of stake (PoS) protocol called Casper, which seeks to solve this issue of efficiency and sustainability. This transition will become increasingly important because of the difficulty bomb mining adjustment that was introduced in 2017 through the Byzantium update. The bomb will make mining Ether more and more difficult until it will eventually become infeasible. This also needs a solution if ETH 2.0 is going to run the world of DeFi going forward.
So what are the advantages and disadvantages of a Proof of Stake blockchain for ETH2.0?
n addition to being inefficient, PoW blockchains are susceptible to market monopolization because they bestow unfair advantages on actors with more resources. This is a problem considering Web 2.0 companies like Amazon, Google, and Apple have tons of money and resources and want to control the internet. DeFi is aiming to limit corporate power and transfer that power back to the public. This is why ETH 2.0 is a powerful hedge to Monopoly power and why whomever builds the killer DeFi apps on its back will likely have valuations that exceed tthe Web 2.0 companies. DeFi companies will actively put those companies out of business by decentralizing how they can do business.
Wealthy and powerful individuals and organizations can afford many faster and more powerful computers which gives them a much higher chance of successfully mining each available token. About 65% of bitcoin mining is done by 5 huge mining groups who could theoretically join together to control more than 50% of the market. This has created a situation where bitcoin and other blockchain networks are not nearly as decentralized as they were originally intended which endangers their independence and utility. Recently this has changed with more miners and more countries now coming on line in 2021.
Proof of Stake first emerged in 2012 as an attempt to solve these issues of efficiency, cost and centralization. In fact, the Ethereum team planned on transitioning to PoS from the very inception of the project, but has only more recently (2020) laid out a plan to do so with Casper.
In a PoS system, the entire mining process is made virtual, removing the need for much expensive and inefficient hardware. The expenditure of energy is replaced by economic incentives and game theory as a means to secure and validate. In PoS, “miners” are replaced by “validators” who all posses tokens that are native to the network. These tokens will be considered gas to run on the ETHER network.
For each proposed block, validators stake a portion of their coins on a block which they think will be a valid addition to the chain. If that block gets appended, it will be added to the chain and any validator who bet on it will be granted an award proportional to their bet. Since there is no block award in a PoS system, validators are rewarded with transaction fees. Right now in January of 2021, transaction fees for certain transactions are quite expensive. This should not shock anyone, because the ETH 2.0 network just turned on, so without a ton of users prices will be initially high until more transactions are made and prices should fall. This can be several thousand times more cost effective and energy efficient than a PoW system. This makes it scaleable so it can compete and eventually replace Paypal or Visa. I think it more likely that ETH 2.0 will be absorbed by these companies to run all their transactions on it. This will give ETH 2.0 the volume it needs to succeed.
What are the some other risks we need to consider?
The final major issue that will be addressed by ETH 2.0 is speed and usability. This is related to but not the same as, the issue of scalability that will be addressed by sharding.
What is sharding? Sharding is a technique in blockchain that seeks to achieve scalability within a blockchain network. The process of sharding seeks to split a blockchain network into separate shards, that contain their own data, separate from other shards.
The introduction of sharding should remove the most significant bottleneck for transaction speed and throughput, but another bottleneck is caused by the Ethereum Virtual Machine (EVM) itself. The EVM is essentially the underlying structure of Ethereum that executes code and allows the network to function correctly. The EVM is responsible for the internal state of the Ethereum network and for all of its computations. The EVM also ensures that the following are accurate: account information such as balances, current gas price, addresses and block information. Furthermore, the machine must keep track of the following components: Block Information, Storage State, Account State and Runtime Environment Information. The Virtual Machine also deals with all Ethereum smart contracts which are written in Solidity/Vyper and compiled into unique EVM bytecode. The contracts are then executed by each node on the network.
Because it is responsible for so many important aspects of the Ethereum network, the speed with which the Virtual Machine is able to perform tasks has a major affect on the overall speed and usability of the network itself. The solution to this bottleneck advanced by ETH 2.0 is called Ewasm. The risk there is that Google developers are involved in this network upgrade. Others are also involved to help mitigate the this risk. Engineers from Mozilla, Microsoft and Apple have been added to make sure no corporate bugs are entered at this change to centralize the blockchain. I just do not trust any of these companies right now.
But if they get this right I view ETH 2.0 like the Panama canal. It is a huge engineering project that will change the world conducts its business globally. This canal reshaped the economic landscape of many states in the USA and with Countries in the Far East.
More than a century ago, the opening of the Panama Canal revolutionized international trade by making it much quicker and easier to travel between the Atlantic and Pacific Oceans. Having easy access to a large number of trading partners is an important determinant of where economic activity is located. It also shaped population growth in the USA.
The Panama Canal was recently widened and reopened for business in 2016 as you can see above. This event coincided with the 100th anniversary of the canal. The expansion, named the “Third Set of Locks Project,” enabled the canal to double its capacity by adding a new traffic lane. This will allow ships that are wider and deeper to travel through the lanes and locks.
I view ETH 2.0 to this event in the Panama canal because the goals are quite similar.
Serenity/ETH2.0 is also under construction to satisfy several more general design goals. Because of the many new and complex systems that will arise from the introduction of Casper and Sharding, one of the main design goals of Eth 2.0 is to reduce network complexity as much as possible, even if this leads to some loss of efficiency. This is possible because the massive increase in throughput and efficiency allowed for by sharding, Casper and Ewasm will outbalance a small reduction in efficiency in order to mitigate the additional complexity necessitated by these same systems. Another major design goal is to use cryptography and design techniques that allow for the participation of a large set of validators, both in total and per transaction. A third goal in the same vein, is to create a system that allows a standard consumer laptop to process and validate an individual shard. Both of these goals are meant to ensure that the network remains secure and decentralized so no single government or CEO can co-opt it.
WILL ETH 2.0 BE PROTECTED FROM A CME OR ELECTROMAGNETIC WAR?
Serenity/ETH 2.0 has two long-term design goals created in anticipation of potential issues that may arise in the future. The first is to design the release in such a way that the network will be able to remain live in the event of a major partition or if a very large percentage of nodes goes offline simultaneously. In his talk at Devcon 2018, Ethereum developer Justin Drake explained how the team wants ETH 2.0 to be able to “survive WW3” or a similar situation in which up to 80 percent of nodes go offline at once.
The developers are thinking about CCP and the USA relationships.
For example, one or more major climate change related disasters could knock out a large portion of nodes within hours, or a major power such as China or the United States could decide to ban Ethereum and other blockchain networks, and try to eliminate the nodes in their country.
ETH 2.0 is being built with geopolitical forethought.
The other contingency that ETH 2.0 anticipates is the advent of quantum computing. There is currently an ongoing tech race between the United States, China, and other major powers and corporations to be the first to invent a production quantum computer. At this point the question is not if, but when. Whenever they do come along, quantum computers will be exponentially more powerful than even the most advanced traditional computers. This will render most existing cyber security, including encryption and blockchain tech, relatively defenseless and easily hack-able. It is therefore important for new and future blockchain networks to do everything possible (within the limits of traditional binary computing) to protect against the eventuality of quantum computers. The final design goal for ETH 2.0, as laid out in the Ethereum github repo is: “to select all components such that they are either quantum secure or can be easily swapped out for quantum secure counterparts when available.
On December 1, 2020 ETH 2.0 went live. It will be released in parts as well. Phase 0 will feature a beacon chain PoS system. It will not yet support DeFi Dapps but will lay the foundation for the following phases. Phase 1 will supports Dapps and introduce shards as data chains. Here is where operability risks begin.
Phase 2 will be fully sharded, will enable state execution and will allow for smart contracts. At this point, ETH 2.0 will have all of the major features discussed above. Phase 3 and beyond will consist of iteration, improvement and additional tech. Once each phase is implemented, ETH 2.0 is expected to have the following features: fully sharded blockchain, pure PoS consensus protocol, faster time to synchronous confirmation of about 8–16 seconds and “economic finality” that can be achieved in 10–20 minutes.
Ethereans want ETH 2.0 to become “The world computer as it’s really meant to be.” Investors in ETHER want it to work as designed……..ideas are great but without execution they are deleted to losses.
Want to earn a safe and easy passive income with your crypto purchases? Then try out Blockfi! You can earn: 6% on your Bitcoin balance and 5.25% Ethereum, 5% Litecoin and 9.3% dollars $USDT