If you are following the series, you will see the story of light stress, which was used in an adaptive fashion by mammals 65 million years ago to survive. What underpins this effect, however, may shock you. It is POMC. It turns out that repeated exposure to low levels of mitochondrial stress, which environmental light induces, and various cytosolic and nuclear responses build resilience against higher levels of stress. This response would have significantly benefited mammals during an extinction-level event. This adaptive response, primarily known as mitohormesis, has been shown to extend health span and/or lifespan in several model organisms (Yun and Finkel, 2014).
What are the consequences here?
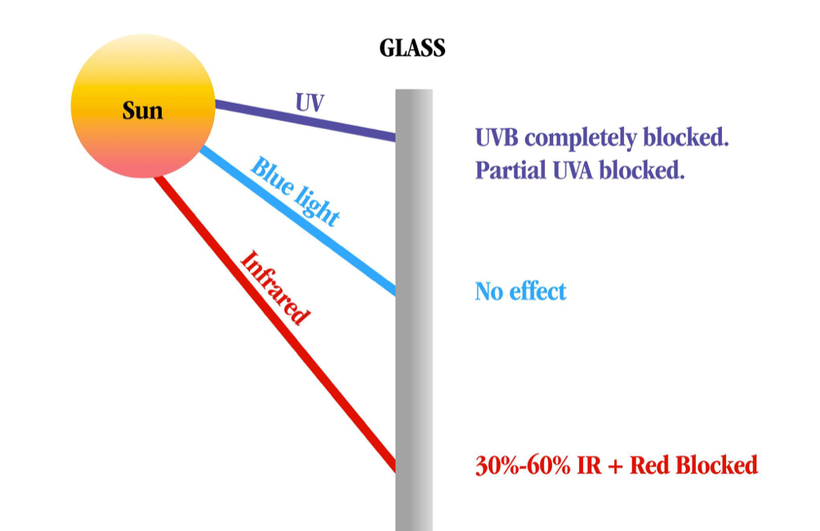
Ancient adaptions can lead to modern diseases when the spectrum of light changes again. Remember, the initial adaptation mammals made was due to a lack of UV light in their environment, and they ran predominantly from 390nm to -3100 nm.
Today, centralized medicine looks at hemochromatosis as a disease when, in reality, it occurred as an epigenetic adaptation for an iron protection strategy that humans used to keep adult males alive to reproduce to allow them to survive in Europe for the last 1000 years. Today, scientists are beginning to realize that our ‘junk DNA’ seems to be the raw material for constructing new wide-bandgap (WBG) semiconductors that use light to sculpt changes. This is how our semiconductors transform light into epigenetic information to change the game so survival is guaranteed. Evolution seems to tell us that in the last 1000 years, biology built a new way to defend against pathogens and events we have recently faced so that we can survive whatever life throws at us. This semiconductive fabrication plant in our bodies (POMC) acts like an evolutionary junkyard that allows us to innovate new novel ways to survive a bad event.
We now know that transgenerational epigenetics in the Viking men of the Northern coastline of Northern Europe was selected for hemochromatosis. We believe that the COLD, harsh Tundra of the north was mineral deficient. Women with this genetic defect would have fared as better child bearers because they could absorb more iron to birth more children who also carried the hemochromatosis defect into the next generation. It is also believed that the Viking men might have survived the disease because their Gladiator-type lifestyle was ferocious, and they often faced severe blood loss that might have offset the iron defect. As Vikings settled in Northern Europe, the mutation grew using the “founders effect” caused by inbreeding due to small population sizes. The founder effect means that any ‘non-lethal defects’ are highly selected for and carried in the entire population of a people. It is believed that the ‘Viking defect’ was blended into the populations of Northern and Western Europe over 500 years to solve the recurrent Yersina outbreaks that caused the bubonic plague and almost extinguished humans in Europe. The chronicity of the infection was an ‘epigenetic signal phenomenon’ that allowed for the selection of the hemochromatosis gene to confer reproductive fitness over longevity. At the same time, the Yersina remained active in the human population. This remained true for close to 500-1000 years.
This “irony” may now explain why ancient physicians were barbers and bloodletters. We used to believe this practice was ‘quackery,’ but we now know that it was the survival of the wisest in action. Bloodletting had a significant role in conferring more longevity to those with hemochromatosis of European descent. When you bleed, you release a massive dose of vasopressin from the posterior pituitary.
Until the 20th century, bloodletting was standard practice. Then, it was stopped, and hemochromatosis became a modern disease. Canadian physiologist Norman Kasting found that bloodletting also released the hormone vasopressin (ADH) from the posterior pituitary. This release from the hypothalamus reduced their fevers and increased their immune function to act faster to save them. This finding was not causation, but the correlation between bloodletting and fever reduction is massive in human history. Bleeding them down may have helped fight infection when it was present. When we bleed, vasopressin release is also altered. nnEMF does the same.
AUTOIMMUNITY AND VASOPRESSIN
Abnormal non-circadian release of vasopressin is linked to autoimmune conditions in modern man.
The arginine vasopressin hormone (AVP) of the posterior pituitary increases blood−brain barrier permeability. It also affects voltage gates and water balance in cells.
The immune system (IS) plays a vital role in protecting our body and can recognize its tissues from foreign molecules. The IS comprises lymphoid organs, cellular components, humoral factors, chemokines, and cytokines that respond to antigens that can harm the body (Shinde and Kurhekar, 2018; Verbsky and Routes, 2018).
The IS is highly and tightly regulated by mtDNA signaling and biophoton biology; however, its disruption could induce diverse pathological disorders, such as allergies and asthma; subsequently, when the host’s tolerance is exceeded, the condition becomes an autoimmune disease (Anaya et al., 2016).
nnEMF exposure is critical to BBB function, as Russ Adey and Allen Frey demonstrated in the 1960s.
Most autoimmune diseases such as rheumatoid arthritis, systemic lupus erythematosus, Hashimoto’s thyroiditis, Crohn’s disease, type 1 diabetes mellitus, and MS are considered chronic illnesses today by centralized medicine. I consider them modern diseases linked to aberrant lighting and nnEMFs.
Some of them can induce neuroinflammatory and neurodegenerative processes in the CNS, which occur when the integrity of the BBB is compromised, as the pictures above show. The BBB is considered a highly selective barrier between the cerebral capillary blood and interstitial fluid of the CNS (Kadry et al., 2020; Schreiner et al., 2022) and helps keep harmful substances from reaching the brain as pathogens, toxins, and some drugs, as well as prevents the entry of IS components (Daneman, 2012; Kadry et al., 2020; Knudsen et al., 2022).
The principal constituent of the BBB is the endothelial cells, which provide protection and structural stability to the blood vessels on tight junctions (the liner sheets pericytes and astrocytes that ensheath the blood vessels and restrict the substances entering the brain or its immune system. The breakdown of the BBB promotes its permeability, permitting the entrance of immune molecules and lymphocytes, inducing autoreactive conditions in the blood-spinal cord barrier and BBB; as a consequence, damage and destruction of the myelin sheath increases, causing neurodegenerative disorders such as Alzheimer’s disease, Parkinson’s disease, ALS, and MS.
Similarly, neuroinflammation promotes the entry of IS components through the BBB and blood-spinal cord barrier into the CNS; therefore, an increase in the barriers’ permeability induces the interaction of pro-inflammatory cytokines such as the interleukins 1β and 17A (IL-1β and IL-17A), and tumor necrosis factor α (TNFα), which activates the downregulation of tight junctions in the endothelial cell barriers. IL-17A has been associated with the loosening of both obstacles, as shown by in vivo and in vitro assays. It is related to the production of reactive oxygen species by nicotinamide adenine dinucleotide phosphate (NADPH) and xanthine oxidase action, which are related to increasing in the endothelial cells’ permeability, causing a decrease in the occluding protein, zonula occludens-1 in in-vitro assays by Arima et al., 2013. The photoswitch I have mentioned before is a critical circadian controller of the ROS mechanism.
In contrast, the hyperactivity of sensitized lymphocytes induces the proliferation and secretion of IL-17 in MS. In addition, under normal conditions, T regulatory (Treg) cells alter and break down the balance in IS responses, which are accompanied by MS (Pot et al., 2011). Immune cells such as T and B lymphocytes, macrophages, and innate immune cells promote the disease’s pathophysiology. Myelin antibodies from B cells induce the loss of the myelin sheath. Furthermore, studies have demonstrated the presence of immunoglobulins IgG and IgM in patients with acute and chronic MS.
THESE DISEASES ARE MULTIFACTORIAL, BUT LIGHT AND POMC DYSREGULATION ALWAYS AT THE CORE
POMC neuronal activity is strongly tied to mitochondrial function. The higher your mitochondrial redox power is on your inner mitochondrial membrane, the more POMC a tissue will express. More POMC = more semiconductors can be made. POMC neuronal activity can be upregulated in mice by mitochondrial-derived ROS (Diano et al., 2011). Moreover, mitochondrial dynamics (i.e., fusion and fission) in POMC neurons are essential for maintaining whole-body energy and glucose homeostasis under altered metabolic conditions (Ramírez et al., 2017; Santoro et al., 2017) in all mammals. Despite the evident importance of mitochondria-originated signals in POMC neurons (Mishra et al., 2014), the details of the underlying mechanisms remain largely unknown in centralized medical research.
MITOCHONDRIAC UNDERSTANDING: SUNLIGHT IS MANDATORY for making water at CCO during the day in mammals. If you do not get enough sun or live at a high latitude inside all day, you need more water to avoid the vasopressin release issued POMC. If we do not get enough sunlight, we’re dehydrated, and then we lose circadian feedback control of vasopressin, and the entire water cycle in our body goes awry. This facilitates the development of autoimmunity.
This is how lousy clock management leads to epigenetic disease by decreasing mitochondrial redox power. This is why big pharma is now pushing the use of anti-vasopressin analogs for MS patients. Understanding POMC is understanding modern human neolithic diseases. Mammals are creatures sculpted by light.
Recent work in this area shows that POMC neurons exhibit a dimorphic (biphasic) response to mitoribosomal stress in a dose-dependent manner; homozygous deletion of Crif1 was detrimental (i.e., severe light stress), whereas heterozygous disruption was beneficial (i.e., mild light stress).
How does IR-A light work in the sun? A biphasic dose response has been frequently observed where low levels of red light have a much better effect on stimulating and repairing tissues than higher levels of light. The so-called Arndt-Schulz curve is commonly used to describe this biphasic dose response. Centralized medicine has no idea how POMC works with light. POMC gets cleaved using the biphasic actions in light. When cleavage is imprecise, disease results. Now you do, too.
Published research has found that low levels of mitoribosomal stress in POMC neurons induced high metabolic turnover and resistance to obesity through cell-non-autonomous mitochondrial stress signaling between the hypothalamus and adipose tissues. That stress signal is VUV light emission in hypothalamic neurons in the leptin-melanocortin pathway. That is how you stay thin. It would be best to renovate the melanin sheets inside your tissues constantly.
In humans, prolonged or repeated cold exposure without surface-level UV light can increase the mass and activity of brown adipose tissue in the neck and supraclavicular regions, as defined by the uptake of glucose, and can improve glucose homeostasis. This is the off switch for ACTH release from POMC mammals used to survive in high latitude and poorly lit cold areas.
POMC neurons control adipose tissuethermogenesis (Dodd et al., 2015). I knew 15 years ago that this discovery, as pictured below, was coming. Brown adipose tissue works its job because of melanin. Surprise!
Mitochondrial communication via optical transmission is the key to adaptive stress response following mitochondrial perturbation. Recently, small functional peptides encoded within the mtDNA, known as mitochondrial-derived peptides (MDPs), have been identified (Reynolds et al., 2020). MDPs represent a unique class of mitochondrial-encoded signaling factors that respond to mitochondrial stress and promote health/longevity (Galluzzi et al., 2018; Mottis et al., 2019; Quirós et al., 2016; Tan and Finkel, 2020).
Notably, MOTS-c (mitochondrial open reading frame of the 12S rRNA-c) mediates mitonuclear communication by translocating to the nucleus upon metabolic stress and regulating adaptive nuclear gene expression to promote cellular homeostasis (Kim et al., 2018).
Light sculpts your life. VUV light, to be exact, when it comes to autoimmune disease conversion. That tells me that mtDNA biophoton release is where the defect is because terrestrial sunlight does not contain these frequencies.
Vasopressin (AVP) is a crucial hormone regulating water balance and is released during hyperosmolality to limit renal excretion. It has a long history to explore. It is produced by adjacent melanin in humans. Arginine-vasopressin (AVP) is a nonapeptide that is synthesized mainly in the supraoptic (SON), paraventricular (PVN), and suprachiasmatic nucleus of the hypothalamus. In addition, AVP is produced in several other brain areas and organs, e.g., the medial amygdala, bed nucleus of stria terminalis (BNST), and the adrenal gland chromaffin cells. Its release is associated with any body stress response, opening the blood barriers to the gut and brain.
SUMMARY
When vasopressin is altered, so is iron biology, which leads to an altered amount of ROS/RNS. Why?
Why do we say oxygen is “reduced” when iron is oxidized in our body?
We say this because iron has gone from its elemental state with no charge ( Fe0) to its ionic state (Fe3+). Because the iron has lost electrons and become positively charged, it has been oxidized. The oxygen has been reduced because it gained the electrons iron donated to it. The electrons from the iron went to the oxygen. Every oxidation process has to have a corresponding reduction.
Iron exists predominantly in two biologically relevant redox states: ferric iron, the oxidized state (Fe3+), and ferrous iron, the reduced state (Fe2+). Fe2+ is well known to facilitate electron transfer reactions that can lead to the generation of reactive oxygen species.
The involvement of singlet oxygen in biophoton emission has implications for our understanding of many diseases like mast cell disease in the skin that links to immune function. Mast cell dysfunction is related to an absence of 1270 nm light in the skin of mammals. Singlet oxygen is known to liberate this frequency of light, as the picture in this blog shows. People with mast cell disorders do not make enough hydrogen peroxide from their mitochondrial respiration. As a result, a comorbid lack of sunlight containing 1270 nm light and lack of H202 creation in tissues is associated with immune dysfunction in mast cells. There is a lesson here that the photoswitch in our cells is teaching us about immune dysregulation.
People with autoimmunity, mastocytosis, and poor wound healing are always deficient in AM sunlight. Why? This is when we get a lot of NIR light with 1270 nm. Early morning sunlight, 6 AM -9 AM, has a relative irradiance with a higher amount of photons in the visible and NIR spectrum than midday exposure (noon). The picture tells why decentralized medicine always recommends AM sunlight. This sun time = TINA = THERE IS NO ALTERNATIVE.
CITES
G.T. Dodd, S. Decherf, K. Loh, S.E. Simonds, F. Wiede, E. Balland, T.L. Merry, H.Münzberg, Z.Y. Zhang, B.B. Kahn, et al. Leptin and insulin act on POMC neurons to promote the browning of white fat.
Alvarez, J. I., Cayrol, R., and Prat, A. (2011). Disruption of central nervous system barriers in multiple sclerosis. Biochim. Biophys. Acta BBA – Mol. Basis Dis. 1812, 252–264. doi: 10.1016/j.bbadis.2010.06.017
Anaya, J.-M., Ramirez-Santana, C., Alzate, M. A., Molano-Gonzalez, N., and Rojas-Villarraga, A. (2016). The autoimmune ecology. Front. Immunol. 7:139. doi: 10.3389/fimmu.2016.00139
Arima, Y., Kamimura, D., Sabharwal, L., Yamada, M., Bando, H., Ogura, H., et al. (2013). Regulation of immune cell infiltration into the CNS by regional neural inputs explained by the gate theory. Med. Inflamm. 2013:898165. doi: 10.1155/2013/898165
Baron, J. L., Madri, J. A., Ruddle, N. H., Hashim, G., and Janeway, C. A. (1993). Surface expression of alpha 4 integrin by CD4 T cells is required for their entry into brain parenchyma. J. Exp. Med. 177, 57–68. doi: 10.1084/jem.177.1.57
Microglial cells declined by 80% in those with cognitive issues, the study found. POMc is located in microglial cells.
RPE cells are located between photoreceptor cells and the choroid membrane, with the basal side connected to Bruch’s membrane and tip microvilli connected to the outer segment of photoreceptor cells (Figure 1).
Damage to the structure and function of the retinal pigment epithelium leads to a variety of retinopathies, and there is currently no curative therapy for these disorders. The reason for this belief is because modern medicine has no idea that renovating POMC internally by improving mitochondrial redox power and optimizing hemoglobin delvery is the key to RPE repair.
Light in our environment plays the most dominent role in obesity. The slide below shows how badly the food gurus have missed the boat.
Within the central nervous system, energy homeostasis is largely controlled by a fine balance between orexigenic and anorexigenic neuropeptides in the hypothalamic arcuate nucleus (ARC). Neuropeptide Y (NPY) and Agouti-related protein (AgRP) are coexpressed in neurons of the ARC and are potent orexigenic peptides, whereas proopiomelanocortin (POMC) precursor protein in the ARC is cleaved into potent anorexigenic α-melanocyte-stimulating hormone peptides. NPY/AgRP and POMC neurons in the ARC are considered “first-order” sensory neurons in the control of energy homeostasis (1) and receive, coordinate, and respond to changes in nutrient and hormonal fluctuations associated with changes in metabolic status. NPY acts on Y1 and Y5 receptors in the paraventricular nucleus of the hypothalamus to stimulate feeding, whereas acetylated α-melanocyte-stimulating hormone and AgRP peptides act as agonists and antagonists, respectively, at the melanocortin 4 receptor (MC4R). The POMC and AgRP interaction at the MC4R is collectively known as the melanocortin circuit. The critical importance of the melanocortin system in food intake and energy balance is highlighted by conditional gene ablation experiments. Ablation of POMC neurons in adulthood produced an increase in food intake and body weight and caused glucose intolerance, whereas ablation of AgRP resulted in rapid hypophagia, weight loss, and starvation = anorexia
Where the defect is on the retina and skin determines whether you get obesity or anorexia. We see this in drug addicts, alcoholics, and we see it in anorexics who live inside behind glass and infront of sceens. Both conditions, however, are associated with altered signaling in the leptin melanocortin pathways.
Why did obesity really happen dramatically after 1874 when the power grid was begun? LIGHT. It brought us INSIDE where the PROBLEM was.
When we started plugging in to the AC power all hell broke loose and this is where MOST chronic disease epidemics began. HARD STOP
Governments allowed NGOs like Gates and dermatologists began to advocate blocking the sun while putting screens in front of millions of humans from 1947 on. In 2009 Barry Obama and Biden changed FCC broadcast transmission from analog to digital and this brought LCD and plasma screens. Now LCD screen make up 99% of screens. All screens default setting out of a FACTORY ARE SET to blue frequencies. THIS MAKES YOU MORE COMPLIANT TO GOVERNMENT PROGRAMMING. The evolutionary time line of light makes this abundantly clear. Food gurus like the MEANS SIBLINGS are like propaganda wings of the Federal government who are now weaponizing MKULTRA programming at GLOBAL SCALE. They are paid by a16z who is a Silicon Valley investment group who supports Big Tech where all blue light begins.
In 2007 Barry Obama said we are also going to remove incandescents which contain some UV and IR. the government knew what they were doing. They knew it make you more susceptible to things coming from Wuhan. They told us that it was to lower energy bills. That was a PR bullshit story that the media sold and then the food gurus were weaponized like legacy media reporters to repeat the liwe so you’d all believe it. AND 99% of you have.
They did it to lower our dopamine levels to make us more responsive to the political moves. This also lowered out melatonin levels simultaneously and this is why all kids now get Rx for melatonin from pediatricians who are complicit in the chronic disease epidemics. When dopamine and melatonin drops SO DOES POMC TRANSLATION. This means melanin never gets made properly. THAT IS WHERE YOUR DISEASE EPIDEMICS COME FROM.
Mental disease, drug addiction and obesity all come from obstruction of this pathway by modern lighting. LOOK AT THE PICK ABOVE. YOU ARE BEING LIED TO IN A BIG WAY by the NGO and Government partners in BigHarma and Big Tech that both sit at the CANTILLON EFFECT. Calley and CASEY MEANS ARE PAID BY THESE PEOPLE and this is why both of them have companies that use WBAN technology which MAKES YOU SICK. They got this idea from the Wojcicki sisters of YT and 23andme and YT and Google’s Sergei Brin. See this here to show the PROOF —–> https://threadreaderapp.com/thread/1826512218088722575.htmlXIA
THE DRUG ADDICT STORY BELOW LINKS TO WHY SOME GET ANOREXIA.
SUMMARY
In the leptin melanocortin system of the retinohypothalmic tracts in the eye the, hormones of the “fed state” such as leptin and insulin, are released in the bloodstream by adipocytes and by the β-cells of the pancreas, respectively, cross the blood-brain barrier to bind to leptin and insulin receptors on the surface of pro-opiomelanocortin (POMC) neurons to promote processing of POMC to the mature hormone α-melanocyte-stimulating hormone (α-MSH), which signals to decrease energy intake.
The three MSH molecules from proopiomelanocortin (POMC) decrease appetite, increase satiety, and increase energy expenditure:
Alpha-melanocyte stimulating hormone (α-MSH)
A POMC-derived neuropeptide that binds to melanocortin-4 receptor (MC4R) in the brain to suppress appetite and increase energy expenditure
Melanocortin-4 receptor (MC4R)
A receptor expressed by neurons in the paraventricular nucleus (PVN) that suppresses food intake when bound to α-MSH
POMC neurons
Neurons in the hypothalamus that produce α-MSH and other peptides that act on MC4R-expressing cells
Leptin and serotonin are signals that promote POMC neuron action, which leads to the production and release of α-MSH. In contrast, ghrelin is an orexigenic hormone that activates AgRP neurons, which antagonize MC4R signaling and increase food intake.
Mutations in the POMC gene have been linked to severe childhood obesity.
STOP LETTING THEM LIE TO YOU.
IT ALL BEGINS WITH LIGHT
CITES
Zarbin M. (2016). Cell-Based Therapy for Degenerative Retinal Disease. Trends Mol. Med.22 (2), 115–134. 10.1016/j.molmed.2015.12.007
Tian X., Cui Z., Liu S., Zhou J., Cui R. (2021). Melanosome Transport and Regulation in Development and Disease. Pharmacol. Ther. 219, 107707. 10.1016/j.pharmthera.2020.107707
Robert O. Becker, MD, found that some part of matter in our nervous system created a DC electric current, and that current drove all healing and regeneration. That current also broke with convention because it could de-differentiate RBCs to primative forms to create stem cells to regenerate our tissues. The current needed was precise. One-thousandth of a million ampere of current was required to do this. Anything less or more than this was a problem. This was the electromagnetic criticality that mammals needed for well-being. When I met Becker in 2007, I told him that POMC was the source of his work. Light was the source of man’s regenerative current. This idea was more heterodox than his journey into bone regeneration.
The phenomenon of criticality has been postulated to explain the sudden emergence of new properties in a wide range of complex systems, from avalanches to flocks of birds to stock market crashes. I believe it is behind the modern epidemics of autism and neurodegeneration, too. I think it is exacerbated by things that reduce our redox potential, like vaccines and modern lighting. Neuroscientists are now seeking evidence that criticality is at work in the brain’s networks of neurons.
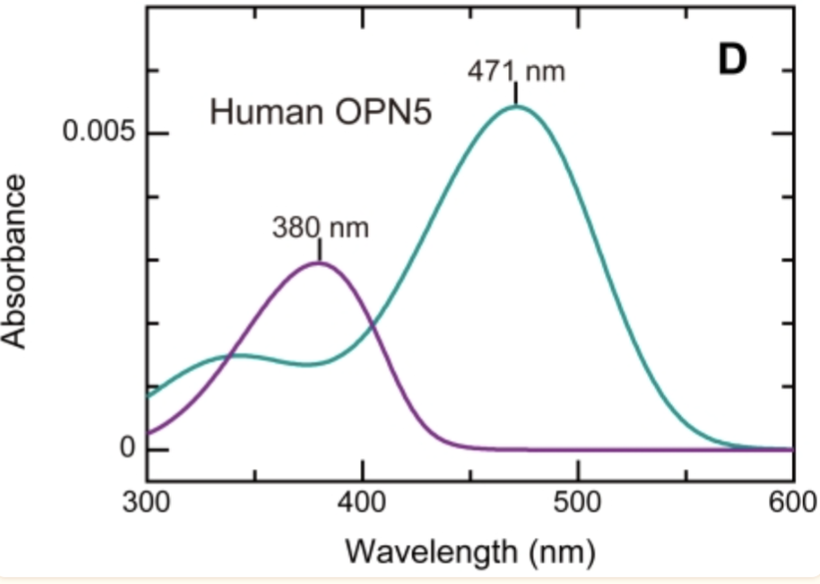
Criticality is linked to how the OPN5 melanopsin receptor works in the human brain. It is the critical opsin that defines modern chronic diseases.
I believe self-organizing criticality theory (SOC) needs to critically look at the work of Ilya Prigogine on how dissipative systems operate. I believe this is why SOC is stuck in the mud with neuroscientists. To understand this idea you need to understand how the liberation of Vitamin A from our non-visual photoreceptors begins the process of diseases by destroying all of our photoreceptors.
People often forget that all of the cytochrome proteins in our mitochondria are also heme-based photoreceptors linked to this damage cascade. This is found in every human disease I have studied, from autism and Alzheimer’s to zoonosis.
All mitochondria liberate heat. and melatonin and water Heat is linked to universality and universality is linked to the fractal design found in nature.
Joseph Fourier made an essential contribution to physics in the nineteenth century when he published his mathematical formulation of how heat propagates. Today, Fourier’s law tells us that heat flows from regions of higher temperature to areas of lower: “that heat flux is proportional to the difference in temperature.” This law is a remarkable insight into universality. Here, we have many disparate macroscopic systems, from solids to gasses to liquids, each containing an inordinate sum of molecules and conforming to a heat conductivity law. To the chemist Ilya Prigogine, Fourier’s description was the “birth of the science of complexity.”
Around the same time, another universality, the first law of thermodynamics, was formalizing in science. It tells us that while “energy cannot be created or destroyed, it can be transferred from one form to another. Energy is forever conserved in the universe.
The critical phenomenon of nature is how light energy is transformed. It can become mechanical, chemical, or vital energy (DC electric current). Light energy undergoes a continual conversion of itself as it interacts with matter. The atomic organization of matter is the key to energy transformation and criticality. This energy attraction occurs throughout space and drives the living force in cells. The energy flow is called kinetic energy, which drives heat into one form of energy or another. Thus, order is maintained in the universe—concerning the total energy in the system, nothing is deranged, and nothing is ever lost. Still, the entire machinery in cells, as complicated as it is, works smoothly and harmoniously.
I believe sleep is designed to get brains back to the SUBcritical state, but electromagnetic signaling from circadian biology is what is required to optimize electromagnetic criticality in neurons. This electromagnetic criticality is linked to the optimized control of ALL charged particles in the entire system. This creates the quantum coherent state in cells. Normally, mitochondria set the system close to the subcritical tone of the system, but light sets off the criticality.
Order Comes Out of Chaos
Everything not alive dies in “heat death”. Order always arises out of environmental chaos. It’s a world described as an engine in which heat is converted into motion only at the price of irreversible waste & useless dissipation.
So energy all goes from one conversion to another and tends toward a final state of thermal equilibrium = ‘heat death.’ Life uses cells to slow this “heat death” by slowing the flow of entropy by arranging atoms ideally in cells. The cell is a dissipative structure that controls the flow of energy so that it can be transformed many more times into useful work. This is the basis of why critical points are needed in Nature’s biological machines inside of cells.
An isolated system is on an inexorable track toward equilibrium, but many interesting things can happen in between. Life at equilibrium is called rigor mortis. Time spent in a far-from-equilibrium state is called life. Health is the slowest form of death we can create with our choices around light.
In health, solar energy is pumped into cells and stored, and entropy decreases. This is the blueprint of how all living dissipative structures operate. The sun is itself a dissipative structure. So is the Parker spiral. It is the heliocentric magnetic sheet that engulfs the entire solar system. This magnetic sheet in the solar wind affects all living things on Earth via their mitochondria. This is partly how our brains remain subcritical at the proper times. Sunlight alters our tissues to induce changes in our charge density. In animals, electrical information generated in mitochondria is more crucial than genes that form tissues, but few people realize this today in centralized science. It turns out that charge density variation by moving electrons around our tissues is the key to understanding what life is up to when it is connected to the decentralized energy systems of Nature. That is how super-criticality is found during the living state.
WHAT DOES LIFE LOOK LIKE WHEN IT RESIDES ABOVE OR BELOW THIS CRITICAL POINT?
Diseases are processes associated with chronic energy loss and entropy gain. The causes can be vast, but the endpoint is always the same.
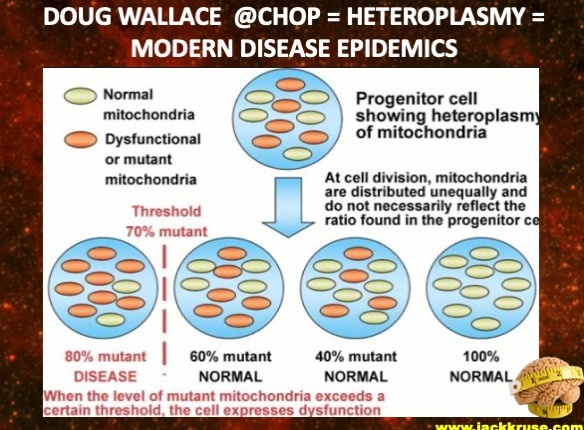
When life is lived below the critical point for any reason, heteroplasmy rates expand in those tissues until energy flow ceases, organ failure takes over, and death is invited to your life.
Earthian life is an open system—doing as much as it can with the solar energy it receives from our aging Sun. The living cell represents an incessant metabolic activity, where thousands of chemical reactions take place simultaneously.
Fasting Factoid: Is fasting tied to a solar mechanism related to iron hemoglobin? Before you answer, think about these facts about fasting. The blood diminishes in volume in proportion to the decrease in the size of the body during a fast so that the relative blood volume remains practically unchanged during a fast. The quality of the blood cells and plasma is not impaired; indeed, an actual rejuvenation of the blood may occur if that blood is allowed to touch the rays of the sun because of how porphyrins work with sunlight.
The old literature on fasting has pointed out that the first effect of a fast is to increase the number of red blood corpuscles. This increase is an acute phase change only and improves the quantum yield if the blood is allowed to be irradiated by sunlight. If the fast goes too long, the spleen acts to decrease the RBC’s mass. When this happens, the astute clinician should expect mitochondria in tissues with high heteroplasmy rates to lose more energy and enlarge as things do when they lose energy (think heart failure or a dying star). The acute increase of erythrocytes during the early part of the fast should be regarded as an improved flow of energy from mitochondria due to autophagy and a cessation of overeating. This red blood cell mass increase has been repeatedly observed in acute anemia cases throughout 20th- 21st century medicine. The decrease in RBC mass is only observed after the starvation period is reached. What are the implications of this, and how might we mito-hack this with new devices that could take advantage of this unique aspect of RBC mass?
When humans are sleep deprived or suffering a circadian mismatch, their brains become supercritical, although a good night’s sleep can move them back toward the critical point. It thus appears that brains naturally incline themselves to operate near this critical point, This points out why sunrise every AM is critical to resetting the critical point in the brain every day. POMC activation is the key to regeneration.
The miracle of our brain’s physiology isn’t that you can see the world as it is. It’s that you can see the world as it isn’t. My job is to explain what you can’t see, but physics has been observed to exist. The absence of evidence in a biology book is not an absence of effect in nature.
The claim that the cortex operates near the critical point is a sweeping one, encompassing optimal information processing, neurological health, and a nearly universal application across species. The need for strong scrutiny is not surprising.
So, why is this view of the critical brain still just a hypothesis? While the evidence in its favor is good, it’s still under discussion.
Early critiques pointed out that proving a network was near the critical point required improved statistical tests. The field responded constructively, and this type of objection is rarely heard these days. More recently, some work has shown that what was previously considered a signature of criticality might also be the result of random processes. Researchers are still investigating that possibility, but many of them have already proposed new criteria for distinguishing between the apparent criticality of random noise and the true criticality of collective interactions among neurons.
Meanwhile, over the past 20 years, research in this area has steadily become more visible. The breadth of methods being used to assess it has also grown. The biggest questions now focus on how operating near the critical point affects cognition and how external inputs can drive a network to move around the critical point. Ideas about criticality have also begun to spread beyond neuroscience.
Citing some of the original papers on criticality in living neural networks, engineers have shown that self-organized networks of atomic switches can be made to operate near the critical point so that they compute many functions optimally. The deep learning community has also begun to study whether operating near the critical point improves artificial neural networks.
SUMMARY
Dreams seem natural when we are in them, but it is when we wake up that they begin to seem strange to us. Might this be evidence of how a point of criticality operates in the mammalian neural network? At the critical point of CNS, fluid acts as a single phase of matter. These fluids are both liquid and gas in quality. These fluids are highly compressible at a critical point, and they allow for travel into other dimensions of the mammalian network. Here are some other significant ideas linked to this story. POMC is required with mtDNA water to create the DC current of Becker. This is the critical point you need to acquire in your journey of wellness.
Sunlight stimulates T lymphocytes in the skin through a mechanism separate from vitamin D production. Sunlight energizes T cells, which play a central role in human immunity. T cells, whether helper or killer types of lymphocytes, need to move to do their work, which is to get to the site of an infection and orchestrate a response. The study below shows that sunlight directly activates critical immune cells by increasing their movement. How do they move? Light in the sun creates H2O2, which makes cells move. This shows that free radical signaling has a positive connotation, not a negative one. This is a magnetochemical signal in the sun’s light.
T cells possess intrinsic sensitivity to blue and UV light. Solar light detection is coupled to the generation of H2O2 and activation of Src kinase and PLC-γ1, leading to elevated intracellular [Ca2+]. Non-visual photosensitivity is greater in activated T cells and enhances T-cell motility.
The majority of T lymphocytes are found in our skin. NK cells come from T lymphocytes. NK cells are cytotoxic lymphocytes that have drawn considerable attention recently as a promising tool for immunotherapy in patients with various refractory hematological malignancies and metastatic solid tumors. I have a sense that the SV40 promotor inactivates T cell motility in turbo cancer formation.
Why? Only a partial response has been shown in centralized clinical results of experimental protocols, attributed mainly to the relatively low number of NK cells infused and their short in vivo persistence. A significant challenge, therefore, in advancing the clinical applicability of NK cells is to expand ex vivo NK cells that display increased functionality upon in vivo infusion. Different combinations of cytokines have been studied to induce NK cell expansion. Sunlight, with fasting, increases NAD+ in cells and is active during the leptin-melanocortin pathway. This might be critical for jabbed patients to avoid oncogenesis due to the presence of 60 billion copies of the SV40 promoter per jab.
SV40 Ori’s normally need T antigen to replicate. This is true unless they have ColE1 and F1 origins, as seen in the Pfizer vaccine.
I also think CBD oil needs to be studied in those who took the jab based on Kevin McKernan’s work and our results with people who used it during COVID-19.
WHY?
A lesson on the quantum biology of photosynthesis.
Did you know that we get a 40% higher yield of cannabis plants at 1200 ppm CO2?
Currently, the biosphere is starved of CO2 at 420 ppm.
The architects of our vaccine bioweapons program support lowering CO2 further.
They also want to block the sun, which would keep CO2 emissions lowered.
Might that be because Cannabis and nicotine were an antidote to the designs they employ? Did you know King Charles just announced a plan to ban nicotine products in the UK by 2030? Do you think he knows something?
Cannabis split from Humulus lupulus (hops) 24M years ago.
Why is its photosynthesis optimized for 1200ppm CO2?
ANSWER: It is because of the effect of deuterium on water. Plants grown in D(2)O show a decreased tendency to fractionate carbon-13 during photosynthetic incorporation of carbon dioxide. The isotopic ratio C(13)/C(12) of the tissues of deuterated plants appears to be proportional to the deuterium content of the tissue. This effect was found in specimens of the partially deuterated vascular plant Nicotiana tabacum, cannabis, and in cultures of the fully deuterated alga Chlorella vulgaris.
Another trick BigHarma plays is using an aluminum adjuvant, which allows you to camouflage all the DNA in your shot. Why? Aluminium is a perfect atomic reflector of UV light biophotons, so it blocks the optical signaling from DNA to the ribosomes and mtDNA to stimulate protein translation. This has a massive effect on POMC translation as a dopant contaminant.
When Gardasil was first approved in 2006, Merck assured the FDA in the USA that there was no HPV DNA in the vaccine. However, this was challenged in 2011 when Sin Hang Lee found HPV DNA in a young, sexually naive girl who had never been exposed to the virus but had received Gardasil. This was due to the atomic effects of contaminant atoms. They ruin the semiconductor of cells and destroy optical signaling, making disease generation more likely.
In 2011, the FDA admitted residual DNA in the vaccine but said it was “expected” to be in products manufactured using recombinant technology and that Gardasil posed “no risk to vaccine recipients.”
The FDA claimed that the presence of DNA fragments was not a problem without showing any studies to prove it had been investigated or that it was safe for humans.
Is there a risk that residual DNA fragments in Gardasil can enter host cells and integrate into the genome? I believe there is now good evidence that this is the case. All one needs is a “transfection agent,” and some studies suggest adjuvants in vaccines can act as transfection agents.
In 1985, the FDA set an upper limit of 10 picograms per dose. In 1987, the WHO increased its recommended limit to 100 picograms and then again to 10 nanograms (i.e., 100 times higher)—a limit now adopted by the FDA.
The paper pictured above argues that it’s difficult to quantify the levels in Gardasil jab, though. HPV DNA is tightly bound to the aluminum adjuvant (AAHS) and forms an insoluble precipitate, which gives it incredible substantivity in a cell and makes genomic intercalation more likely.
Most genomic experts can easily detect the HPV L1 gene DNA in the insoluble precipitate and the soluble DNA in the solution using nested PCR with Sanger sequencing for confirmation.
Genomics expert Kevin McKernan, the first to discover residual DNA in Pfizer’s COVID-19 vaccine, taught me this lesson during our collaborations. He agrees with Dr. Lee’s papers above that the FDA’s permissible limit of 10 nanograms is futile because it increases the cancer risk for the compliant.
Kevin has repeatedly pointed out on his X page that the trick the FDA plays with the guidelines is when you ask scientists to measure the residual DNA, you’ll miss most of the DNA because it is all bound up to the aluminum adjuvant. This is why they add Aluminum to the jab to hide the contamination. This is why one should never comply with any jab. All jabs carry this risk now.
The 10-nanogram limit they’ve created is just smoke and mirrors because they have legal protection now. They say if it’s below that, then they don’t care, but here you have something that hides the DNA in aluminum, and they whistle past the graveyard of patients who took their shots.
Aluminum is one problem, but now mRNA vaccines are known to have 55 different atoms present and SV40 promoters, all of which can perform this task. I also believe mRNA technology in food will pose this EXACT risk to jabbed people.
SUMMARY
I have spoken with Dr. Alexis Cowan about how sunlight, specifically UV light, operates the wiring diagram of the mitochondria. You can find that here. It essentially shuts it down while increasing skin T lymphocytes to become NK to hunt for cancer cells—people who took the jab need to upregulate their own NK T lymphocyte production. Sunlight with UV light appears to be the best way to do this. It should be the key option discussed with patients to avoid oncogenesis.
In decentralized medical research, researchers interested in using exogenous nicotinamide (NAM) supplements to help the jabbed should be considered in those who took jabs known to have the SV40 promotor. I plan on discussing this further with Nicole Shanahan soon after the election. We need this study for those who complied and took the jab without knowing their risks. The slide below shows why I think 24/7 exposure to daily 380 nm light is the best plan for the jabbed. It directly helps metabolism, but it actually repairs the circadian clock gene dysfunction—the jabs also CAUSE circadian disruption.
NAM is a form of Vitamin B3 and a potent inhibitor of enzymes that use NAD+ for their activity. Hence, NAM might be directly involved in controlling redox-sensitive enzymes, mitochondrial functions, cell metabolism, energy production, and cell motility. The studies done on NAM by Josh Rabinowitz and Charles Brenner have not been kind to NAM or their analogs, but in post-jabbed patients, the results may be quite different, and I believe this should be studied.
My perspective on chronic disease creation caused by the jabs is a different perspective for disease modeling than exists today in centralized science. It is far more coherent and discernible to understand when you examine the simplicity of the idea.
Biologic information from the environment must have a connection to its source in cells; that source is mtDNA. The mtDNA then makes a wireless connection using light to the sun and Earth, which is paramount for wellness. Only then can the data from nature remain reliable and pliable for life’s uses through all these physical laws I have given you over 15 years.
Only living things sense and seem to know about their connection to “ex-formation” by the mechanisms built into how mitochondria can feel our connection to Mother Nature. That connection is hard-wired into every cell by its mitochondrial genome and copied and pasted into the maternal germ line of all the genomes of life.
In this way, mothers must remain connected to nature to pass information on accurately. The disease can manifest in children when mothers do this. The child does not need to sense this to get a disease because all mitochondrial DNA comes from our maternal line. This is childhood cancer, depression, and suicides have been on the rise since 1900. No one sees what I see, but they will because of what happened with COVID jabs. This is why the jab architects now weaponize the Means siblings on media. They want to rewrite jab history like Bernays taught the Industrial-military complex.
Mitochondria with high heteroplasmy should never be connected with an excellent nuclear genome if one wants to stay healthy. This is obvious during pregnancy when making a child, as decentralized 16 explains. Leptin resistance in a mother is a sign of avoiding pregnancy; it blocks fecundity. It is also why infertility rates are skyrocketing in the Western world, where technology use is rising, ala MKULTA/Brain Health Initiative. The kids born to these parents are now facing mitochondrial diseases like cancer, diabetes, depression, and suicide at unparalleled rates.
This is akin to putting an Apple computer into a Windows server. Putting insufficient data from mitochondrial DNA into nuclear genomes that are not coherent leads to all disease categories. That is, conditions of existence and natural selection are working in order. They are the Ying and Yang of epigenetics.
Centralized science of vaccination and light use is now genetically engineering changes to the mitochondrial DNA using excessive blue light and nnEMF to expand their distance, ruining our cell’s data processing ability. In this sense, our modern civilization is genetically engineering us to extinction, and the biological information cells continue to receive will get more and more misunderstood until the genome responds with nonsense code. Nonsense code = Chronic disease illness.
The best example of nuclear genome nonsense coding = is cancer. This is how the expansion of the cytochrome proteins over time can lead our cells to oncogenic change. The implication: Might mutations in mitochondria be needed and required from an evolutionary perspective to get us to change what we are doing to remain healthy? Yes, that is my answer. Thus, the most helpful answer for life today is to stay in direct contact with its “ex-formation” to avoid this situation where the disease has to warn us to change our environment. This is why wild animals migrate. They are in tune with nature because they are appropriately connected to the sun and Earth. They do not live indoors as a man does and take BigHarma jabs. The light that regenerates you is the answer. It is never food first unless you are working for the opposition. When you know better, you simply do better.
THE TAKE AWAY:
Electrons are like glue to light as words are to their meaning. Electrons can catch terrestrial sunlight, like words, and capture meaning to create the world we experience. We need sunlight to produce NK killer cells after SV-induced cancers. We can only hope that our words can capture what we mean, but we know they can’t possibly capture that much joy, grief, or wonder…….or can they? I’ve been telling you ALL for some time to spend less time with food gurus like Calley and Cassie Means and way more time with bio-physics because that is where the treasure is buried. I believe the Means siblings are nefarious in this game as well.
Those are the globalists who are all in on the global vaccination plan I warned about in the Danny Jones podcast.
EXPLAIN centralized PEER REVIEW link to the MOSSAD and the US Federal government LIKE YOU ARE THIRD GRADE
I’ll repeat it… “Peer review” is a method used in centralized science to strip you of individual medical sovereignty.
If you outsource your thinking to centralized experts who are controlled by the BigHarma Gutenberg printing press, you will wind up being chronically wallet-biopsied by the industrial healthcare complex. Do you understand the game yet?
Remember that the PEER review is run by dual passport holder (UK/Israel) Robert Maxwell. His daughter is in jail protecting the Zionist Federation guys and the Royal Family. Maxwell was a primary actor in the MEGA Group. She worked for Epstein, and the MEGA Group employed Epstein.
Epstein has ties to Israeli intelligence and well-documented ties to influential Israeli politicians and the Mega Group. Those guys are layered links to the MOSSAD and Meyer Lansky.
Ronald Lauder, a cosmetic heir who makes billions blocking women from the sun, is a Mega Group member. He has known Trump for close to 60 years because they are both Wharton Alumni in Pennsylvania. He is a former member of the Reagan administration and a long-time donor to Israeli Prime Minister Benjamin Netanyahu. He is very influential in Israel’s Likud Party. He was a long-time friend of lawyer Roy Cohn. Robert Kennedy Sr. was Cohn’s assistant to Senator Joe McCarthy in the 1950s during Eisenhower’s administration. Cohn’s father ran B’nai B’rith as its President. Roy Cohn was one of the first group of Americans who received the Salk vaccine, and he died of AIDS in 1986. Sadly, no one ever checked him for SV40, but if I were in power, I think I’d look at his remains and sample them for SV40.
Lansky and his friends in MEGA lured Cohn to NYC to work for the accounts of MURDER Inc in 1954 and 1955, Cohn reportedly had deep connections in the 1950-80s to Ira Malnick.
Lansky worked on cryptography from 1969 to 1983. That work linked art sales and money laundering through the IRS and Treasury via infiltration of the Executive branch. This work created the PROMIS software program. It was stolen in a bankruptcy conspiracy by the DOJ and Industrial military complex corporations, Wackenhut. This program allowed Lansky and his associates to follow the Fed, Treasury, and IRS’s actions while also monitoring Israel’s financial movements. This gave Lansky data on all their financial interactions to see how well their programmable money project was progressing. I covered this here: https://kruselongevitycenter.com/djonesqapublic
This code in this software had dual use that allowed the US and Israeli governments to launder money for their intelligence arms. It also allowed Lansky and his syndicate to stay ahead of his partners in the Industrial-military complex by following where money was being moved. This software was critical in setting up the money laundering schemes from cartel smuggling and set the stage for the Iran Contra money laundering schemes that linked the MOSSAD to the Executive Branch. It also linked the drug trade into Mena, Arkansas, where John Gotti and the governor made sure that the project operated undisturbed.
These are the people I think Calley and Casey Means are working for.
If you understand the science of nnEMF you’d understand why the picture above is an oxymoron. The number one indication for spinal cord stimulation in the USA, according to CMS data, is diabetic peripheral neuropathy (DPM)
The global spinal cord stimulation market size was valued at USD 2.88 billion in 2019 and USD 2.41 billion in 2020 and is projected to grow USD 4.12 billion by 2027, exhibiting a CAGR of 8.0%. North America dominated the global market with a share of 73.96% in 2023.
DO SPINAL CORD STIMULATORS MAKE DECENTRALIZED SENSE FOR CHRONIC PAIN for DPN?
Is “the juice worth the squeeze? Spinal cord stimulators were conceptualized based on the overly simplistic “gate control theory of pain” proposed by Melzack and Wall. Wall himself later wrote, “The least, and perhaps the best, that can be said for the 1965 paper is that it provoked discussion and experiment.” Is that still true based upon this paper written 10 years ago at the 50th anniversary of this “theory.”
HAS IT REALLY REVOLUTIONIZED OUR UNDERSTANDING OF PAIN, OR WAS IT USED TO BLOCK US FROM THE TRUTH ABOUT HOW SUNLIGHT REDUCES PAIN AND DIABETES?
The 1965 gate control theory of pain describes how non-painful sensations can override and reduce painful sensations. A painful, nociceptive stimulus stimulates primary afferent fibers and travels to the brain via transmission cells. Increasing activity of the transmission cells results in increased perceived pain.
So the answer should be a clear no, but we still do them in centralized medicine and CMS data says the market is close to 3 billion dollars. That is a lot of cash for a theory that still has not been proven.
Many people get stimulators for diabetic peripheral neuropathy. How many pain specialists or neurosurgeons know about the links between POMC and neuropathy?
Very few. Why? In 1965, two authors, both linked to academic centers with military funding, proposed a new theory about how pain happens.
Should neurosurgeons and pain specialists know that POMC is critical in DPN and chronic pain? Why aren’t they taught the truth?
POMC PAIN STORY IS A STORY ABOUT LIGHT = MKULTRA STORY
The endogenous opioid system is our body’s endogenous line of defense against noxious stimuli. This system is part of the leptin-melanocortin pathway in mammals. Endogenous opioid precursors are proopiomelanocortin (POMC), pro-dynorphin (PDYN), and pro-enkephalin (PENK)) which are proteolytically cleaved to produce opioid peptides.
These bind to their cognate opioid receptors and trigger downstream signaling events. Those ion channels are activated by K+ channels and inhibited by Ca++ channels. Ironically we know that light and electricity can alter these ion channels. Blu light and nnEMF were known to cause calcium efflux issues in the 1970s. When K+ channels are activated or Ca2+ inhibited is usually part of the clinical event one should expect. These set of circumstances result ultimately in the inhibition of neuronal excitability. Persistent neuropathic pain in DPN = diabetic peripheral neuropathy patients suggests a dysfunction in endogenous opioid-mediated antinociceptive mechanisms. Blue light exposure is known to drive blood glucose high and drive insulin higher. See Nora Volkow’s papers from NYU circa 2011. There is a powerful correlation of blue light exposure to diabetic transformation. Blue light exposure also seems to destroy POMC translation in mammals.
DID YOU KNOW RED LIGHT BY ITSELF LOWERS BLOOD GLUCOSE?
The interesting thing is no one has done a study yet to show that the combination use of UV and IR light might work to reduce pain in the centralized literature.
But did you know that sunlight also reduces the need for insulin for diabetic mammals? It is true, sunlight reduces insulin resistance in all mammals so far tested.
Previous studies have examined the link between the endogenous opioid system and diabetes, and reported that opioid peptide levels are altered in the central nervous system and plasma of diabetic rodent models as well as of diabetic human patients. Today, we now know the opioid levels linked to POMC translation in the peripheral nervous system (PNS) are altered during diabetes.
In fact, in vivo correction of this deficit can rescue diabetes-induced hyperalgesia and associated behavioral changes, thereby showing the relevance of the dysfunctional POMC-MOR signaling to the increased pain sensitivity observed in diabetes. So, should DC electric pain stimulators be replaced with light therapy stimulators since POMC responds to UV light? FYI: Your mitochondria is the UV light generators gone bad in diabetes. This is why all living cells have been shown to liberate ultraweak UV biophotons. Diabetics are no longer capable of this due to their defective mitochondria.
They do not want you to know this because they’d lose 3 billion in generator sales.
GOT IT?
It is well known that UV and IR light reduce blood glucose and decrease pain. Moreover, it is also well known that UV light raises vitamin D levels, and Vitamin D levels highly correlate with increased pain and higher narcotic use. Might light stimulators be better than spinal stimulators because the gate theory idea proposed by science linked to the industrial and military complex is entirely flawed? Better yet, was it really flawed or was it put forth by design by the industrial military complex who paid for Melzack and Walls paper 60 years ago?
Your cells capture the light you choose to live under. This was the message distilled from MKULTRA program. Your choices are quantized in your melatonin and insulin levels. You are essentially an electromagnet for light. As a result of electromagnetic capture, the thoughts created in your brain via your retina and skin have a frequency associated with them and that frequency resonates with a select few. That resonance determines whom you collect in your lives. They are essentially antennas in your life and they are linked by the light you choose.
The centralized paradigm in power in healthcare wants people to think it’s a type of “lock and key mechanism” but this is factually incorrect. It is more like the electro-magnetic gate control we see in semiconducting light diodes. Melatonin receptors are found to a great degree in the retina, RPE, and the visual pathways of the brain. When light is absent then melatonin activates these “light diode-like” gates. For humans, the absence of light is an optical non-linear signal that indicates that it is the period when redox levels are unstressed by solar radiation. This is how the brain tells time, night from day. Melatonin is an old evolutionary clock that was important before the optical lattice clock that now exists in your visual pathways came along via evolution. Light activation can long before food activation in evolutionary timescales.
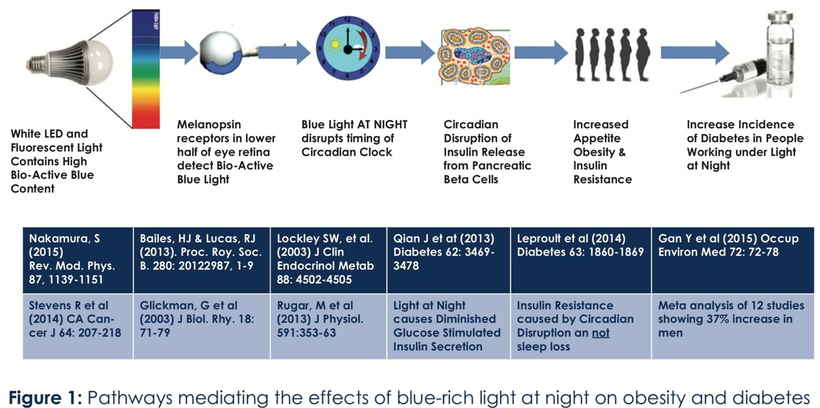
The circadian clock in the eye and tissues, however, in humans still controls glucose and insulin metabolism, as the picture above shows. It also is the reason why blue light at night is what is driving insulin resistance and the runaway diabetes we see today. The optical signal of melatonin is a key circadian timing signal that contributes to and is a part of a cascade of other responses that help initiate and maintain sleep when light is not present. Melatonin levels also control the distances between respiratory proteins in our mitochondria. If the level is low anywhere in the system % heteroplasmy rises and the ATPase of that mitochondria begins to spin slower than it should and electron chain transport slows in that tissue and damage occurs that can make that organ disease. As a result NAD+ drop. This is a result of light not food.
Strangely enough, researchers now know that these receptors are also found in the pancreas, but clinicians have no idea this is what is behind the diabetes epidemic globally. It is related to the amount of deuterium on receptors on alpha and beta cells in the pancreas and a lack of sunlight makes it more likely that you’ll get diabetes. Melatonin receptor density is brisk in pancreatic alpha and beta cells. Melatonin controls the mtDNA in the alpha and beta cells in the pancreas. This is how diabetes manifests from getting too much artificial light at night via our eyes or skin. Most people know beta cells release insulin during the day when we are supposed to eat. Beta cells regulate glucose levels in the blood. During daylight blood pools to the surface to absorb specific frequencies of sunlight while releasing nitric oxide. Researchers also know that when melatonin activates these receptors via the blood plasma, insulin secretion is decreased from the pancreas. This mechanism shows your that diabetes is a photo-electric disease. Your government is behind the epidemic.
The future of maternal fetal medicine is understanding howthe governments MKULTRA programming has ruined the fundamental process.
Did you know the Leptin melanocortin pathway controls the entire physiology of the human placenta? This is why Leptin controls fecundity and fertility. The placenta mtDNA liberates more ultra-weak biophotons, which alter the hydrogen binding networks in the water in blood, and this liquid crystalline structure delivers its payload to the mother brain to begin translation of more POMC deposits in her brain. As water content increases in her circulatory system, so does melanin; this raises her alpha MSH level and increases melanin production in the brain. As a result of the relative rise of UV-powered water in her blood and increased melanin, her brain shrinks. This mimics the process I explained in my recent
Palestra Society talk in El Salvador last month. This phenomenon has allowed me to use cerebral brain volumes on my patients to predict transgenerational risks of the unborn child. This is how the future of maternal-fetal medicine will be practiced in decentralized medicine systems.
Nature always favors the living mavericks in her domain who challenge her rules. She will respond by fighting the maverick all the way, but she respects them and adapts to the environmental pressures the maverick applies. Ultimately, nothing else matters but the pressures placed, which cause her adaptations to occur in cells. They prove that she listens to them much more than Darwin’s ideas in centralized texts.
Morphogenesis develops during the first 5-6 months of pregnancy while the brain remains immature and devoid of development. Suddenly, the brain stops developing in six months, and the child gains more weight than at any other time of its in-utero life. How does this happen? The fetal mitochondria create light spectrum where U light is subtracted with IR light, and blue light is emitted. This fattens the child using melanopsin as its photoswitch. A LACK of endocabbinoidal function turns off the UV 380 nm mTOR photoswitch, and the IR 1280 nm photoswitch is also turned off.
This stunts brain development and fattens the child because there is no POMC translation; it results in a lack of melanin and endogenous endocannabinoid signaling. The green rectangle in the slide below shows you its dynamic effect. As leptin, adiponectin, and endocannabinoids drop, so does mTOR signaling. This means they must affect production of UV biophotons and they have to involved the mix of H+/D levels in the mitochondrial matrix.
Many forget that humans arrest neural development to be able to fit their offspring out of the vaginal canal safely. As neural development halts in humans, the child subcutaneous space needs to fattens up. That will facilitate the child’s passage through a tight canal in the mother, and the fat can be used to mature the brain when the child is born and is put in sunlight. 380 nm light not only matures the child’s brain post natally but also drives regenerative programming everywhere in humans. In fact, endogenous endocannabinoid signaling is critical in regulating decidual senescence and parturition timing.
380 nm light increase cognition in the developing brain. because it is associated with energy flow from matter to be turned on as the slide above shows. Synaptic proteins are upregulated also because of 380 nm because it increases nitric oxide (NO). People forget Nitric oxide (NO) works as a retrograde neurotransmitter in synapses. In brain it does different things than it does outside the CNS. NO allows the brain to increase blood flow to increase energy production and because of this it plays an important role in the fetal intracellular signaling in neurons to regulate the neuronal metabolic status to the dendritic spine growth. Remember as a free radical it controls the magnetochemistry of the fetal brain.
BPA increases deuterium absorption and reduces H+ flow in colony of mtDNA fetal brain. Deuterium in mtDNA is known to cause methylation defects, which causes endocrine dysfunction. Prenatal exposure to BPA at low levels can also affect gene expression in developing brain by turning off growth leading to cognitive decline. Fluoride does the same by stealing electrons that have these light frequencies in the UV range and this limits semiconduction and growth of the immature brain as it develops.
HOW DOES MOM’S PLACENTA FATTEN HER BABY PROPERLY?
Did you know the germinal matrix center in the human brain is found in the area of the brain called the thalamus? The thalmus is where all 5 of our sense target. I told you in the Quantum engineering series that all life has a frequency modulated antenna system built into the thalamus to tell time. Specifically it relates to telling seasonal time. It operates by varying the amount of deuterium in this part of the brain to alter its stiffness. Why?
This systemis built to vary its”flexibility” when it is encumbered by too much mass in the antennae of the system. This is where the Kruse for Dummies lecture comes to into play to explain to you how ATAVISM happens in utero to stop brain growth. It is instructive of why obesity in the adult form is a predictor for nnEMF risk. Humans are designed to fatten when they add deuterium to their thalamus to affect of the organism accounts for timing.
What bends space/time in the universe? The masses associated with the matter in the universe do. As result, physics teaches us that magnetic fields tend to flatten and stiffen the fabric of space-time when masses are added to them and this alters our perception of timing. So adding deuterium to the mitochondria of the developing thalamus stops neurogenesis in utero. The added deuterium alters our perception of the seasons, and this fattens the child in the third trimester. The process also halts melanin by turning off UV biophoton production in the placenta. This limits the neuroplasticity in the neural crest cells in different parts of the CNS of the fetus. If you remember from Becker’s work he said he found that the semiconductive pathway in neurons that controlled regeneration seems to come from just below the myelin level. This area is where POMC is normally found in humans and when UV biophotons are present would cause POMC translation to make melanin to charge separate water to make a ton of electrons that would signal a semiconductive circuit. When you turn off UV biophotons creation from mtDNA in the placenta and in the germinal matrix center you affectively stop brain growth.
Anything that has more mass than the smallest atom used inside of our molecular clocks changes how we experience time. This is AXIOMATIC. Mass bends space/time! As a result, we should expect the Fo head of the ATP to stop spinning as much. In fact, ATP production should slow down. How does this happen in humans. The mother’s placenta make a huge amount of NO to inhibit the fetal ATPase. This means that the magnetic fields created inside the childs developing neurons is effectively arrested. Anytime humans are using the TCA cycle, they are creating the largest magnetic fields in mtDNA.
Now ask yourself what magnetic fields do to space-time based on this picture below. It orders atoms around it to allow them to direct energy flow to create life. This process slows light down in tissues and causes time perception to change. The collapse of the magnetic field by the mother’s placenta halts this process. Centralized OB/GYN has never understood why the human placenta normally enhances physiological ROS production during pregnancy. Now you know why. It is not a bug in the design, it is a design feature to stop the childs brain growth.
What does this concept imply? Time slows as one approaches the speed of light. At the speed of light, time ceases to change because it contains all change.
A mother that does not get enough sun on her abdomen during pregnancy runs the risk of developing preeclampsia (PE). PE pathophysiology result from abnormal placentation due to a defective trophoblastic invasion and an impaired remodeling of uterine spiral arteries, leading to a poor adaptation of utero-placental circulation. Leptin and melanin control this process. This can alter how the brain develops in her child. This is really how autism begins in humans. It is also how many childhood cancers begin and I believe it is where Angelman’s Sundrome really comes from. Altered circadian biology of the mother’s placenta combined with a altered germline in eggs and sperm.
Note on the slide above that matenral levels of Vitamin D links to NO production of the placenta. This is why maternal Vitamin D levels are a huge predictor of problems in decentralized medicine. Mother’s have to use their stored light to turn off the magnetic fields in their child. People have totally forgotten the basics. Nitric Oxide turns off CCO and ATP production as the slide above shows. NO enhances blood flow through the placenta but it turns off energy flow to control the child’s brain growth. Fetal RBC contain mitochondria to make them less able to tell time so the child does not get involved with the mother’s placental signal to light. So having a mitochondria in your RBCs means you are less of a sensor to the SCN in the retinohypothalmic tract. This means the placenta of man controls the timing mechanisms in the baby when it makes sense to turn off brain development.
It implies that the your skin and eye take over this process when you leave the uterus as well.
WHERE DOES THE METHYLATION PROBLEMS COME FROM?
Tetrahydrobiopterin is a cofactor of the three aromatic amino acid hydroxylase enzymes that I have showed you hundred fo times in the slide below. THB is also used in the degradation of amino acid phenylalanine and in the biosynthesis of the neurotransmitters serotonin (5-hydroxytryptamine, 5-HT), melatonin, dopamine, norepinephrine (noradrenaline), epinephrine (adrenaline), and is a cofactor for the production of nitric oxide (NO) by the nitric oxide synthases.
I just told you the placenta controls NO production, didn’t I. Have you forgotten how NO controls methylation production? An altered antioxidant capacity from the mother’s circadian defects alteres her NO bioavailability in her placenta. Without NO can you collapse the ATPAse? If you cannot do this, can you control morphogenesis or organogenesis in the fetus? NOPE.
Altered NO levels leads to methylation problems in the nucleus. Centralized science has forgotten the basics. Why is this true? Altered methyaltion results in part from the reaction of NO with the radical anion superoxide (O2•−), which produces peroxynitrite ONOO-,. This is a powerful pro-oxidant and inflammatory agent. Another mechanism is the progressive inhibition of the placental endothelial nitric oxide synthase (eNOS) by oxidative stress. DIdn’t I tell you over a decade ago in the Holy Trinity blog that eNOS is a key circadian switch in the SCN of all mammals? I did. What happens when you affect it? An altered eNOS results uncoupling viaseveral events such as a depletion of the eNOS substrate L-arginine due to increased arginase activity, an oxidation of the eNOS cofactor tetrahydrobiopterin (BH4). It also implies an alteration in eNOS post-translational modifications (for instance by S-glutathionylation). This is how childhood disease manifest in utero. This is how a child is born with a large amount of heteroplasmy when the system signals are not well controlled by sunlight.
More heteroplasmy = higher disease burden in a fetus even before it has taken a breath.
How big a miss is this decentralized science for every centralized OB/GYN?
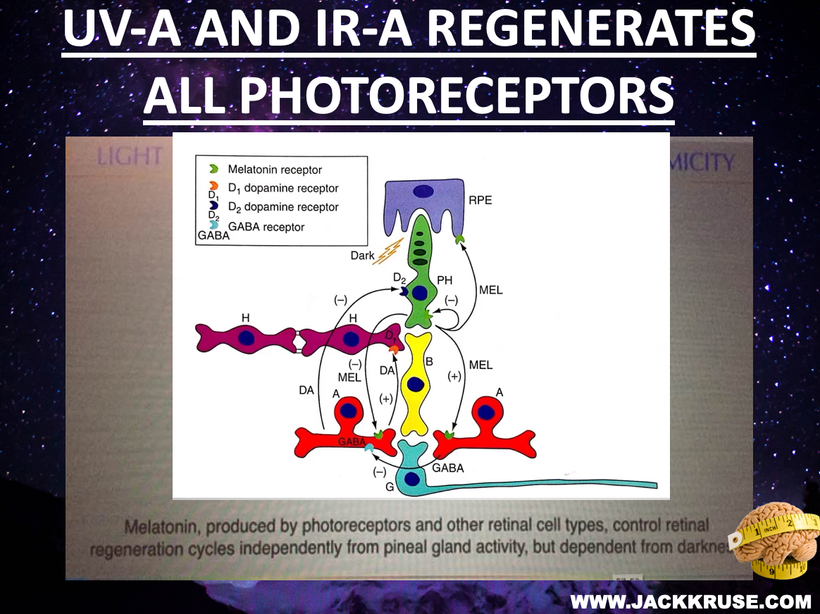
Did you know that tetrahydrobiopterin is a cofactor for tryptophan hydroxylase (TPH) for the conversion of L-tryptophan (TRP) to 5-hydroxytryptophan (5-HTP)? Tyrosine hydroxylase (TH) catalyses the conversion of L-tyrosine to L-DOPA (DOPA), which is the precursor for dopamine. You do remember that dopamine and melantonin control ALL RENOVATIONS OF PHOTORECEPTORS RIGHT? How many times have you seen this slide as proof I have been telling you this?
Now what are all those photoreceptors I keep mentioning? Remember how many times you have seen this slide in blogs too. The same message has been being pound into your blue light toxic brains for decades.
The control of THB and NO by the placenta turns out to be critical in melanin production as the slide below shows. This is the real reason why most centralized OB/GYNs have to give women prenatal vitamins with folic acid. WHY?
BH4 can be oxidized by one or two electron reactions, to generate BH4 or BH3 radical and BH2, respectively. Research shows that ascorbic acid (also known as vitamin C) can reduce BH3 radical into BH4 preventing the BH3 radical from reacting with other free radicals like superoxide and peroxynitrite specifically. Without this recycling process, uncoupling of the endothelial nitric oxide synthase (eNOS) enzyme and reduced bioavailability of the vasodilatornitric oxide occur, creating a form of endothelial dysfunction. This is the only thing centralized OB/GYNs got right. They realized that a lack of folic acid was linked to caudal regression syndromes in fetal medicine. The problem is they still have no idea it links to just about every other disease children get from solar deficit mothers. Ascorbic acid is oxidized to dehydroascorbic acid during this process, although it can be recycled back to ascorbic acid to be reused. Look at the slide above. You’ll see how the placenta turns off the recycling of Vitamin C and now you’ll see why. Glutamate release without Vitamin C turns off neurulation in humans.
Folic acid and its metabolites seem to be particularly important in the recycling of BH4 and NOS coupling. This is why this series has a blog on this. When mothers’ are solar deficient they will need folic acid to prevent many diseases in maternal fetal medicine.
I know you’ve seen this slide below a lot too., but I need to you to connect the lessons I am stacking for you now because centralized medicine is being sculpted by BigHarma to make sure you do not get this lesson via how the curriculums in med school have been hijacked by data that came from MKUTLRA. Blocking this pathway on the top line of the slide below stops brain growth in the fetus!!! It also stops T3 production which is needed for neuron growth and sprouting in humans.
What else is caused by this placental light dysfunction story I am weaving for you?
Phenylalanine hydroxylase (PAH) catalyses the conversion of L-phenylalanine(PHE) to L-tyrosine (TYR). Therefore, a deficiency in tetrahydrobiopterin can cause a toxic buildup of L-phenylalanine, which manifests as the severe neurological issues seen in phenylketonuria. So PKU is a risk factor for women who are solar deficient. I bet your doctor never told you that!
HOW WE CREATE DIABETICS?
The role of BH4 in this enzymatic process is so critical in arresting fetal brain growth that most have forgotten that a deficiency of BH4 in the placenta means – WE SHOULD expect a reduction of nitric oxide production. This is the photoswitch that controls this process. Without BH4 you cannot make NO at all. Recall that Nitric oxide synthase (NOS) catalyses the conversion of a guanidino nitrogen of L-arginine (L-Arg) to nitric oxide (NO).
Why does maternal fetal NO causes gestational diabetes? It should be obvious now. Moreover, it should be obvious why gestational diabetes means the mother and baby are going to be future diabetics. Why do I say this? 30 years of maternal fetal medicine research has pointed to a deficiency of BH4 – and thus, of nitric oxide – as being a core cause of the neurovascular dysfunction that is the hallmark of circulation-related diseases such as diabetes. Anyone who is solar deficient and gets more ALAN is guarranteed to get diabetes because of a chronic lack of NO production. Moreover, why do diabetics get more cancer? NO controls the stem cell depot biology. And a lack of BH4 also causes DNA methyaltion defects which make oncogenesis more likely. EVERYTHING FITS DECENTRALIZED medicine theory here, doesn’t it?
IF YOU BLOCK THE SUN YOU WILL GET CHRONIC DISEASES IN EPIDEMIC FASHION.
I hope youre beginning to understand how atavistic effects predict the future of mankind now as they built a tech world with MKULTA light.
Your mother’s placenta uses light message from her mtDNA to create a defect in the antenna buried in your developing thalamus where all human neurogensis occurs. This optical lattice clock switch in her placental ruins the fidelity of the system in the fetal brain by design so it can no longer tell where the Earth and sun are in relation to one another. It destroys the ultradian ryhythms in the child by design.
FOOD mom eats has zero to do with this mechanism. Her SCN sends this signal to her mtDNA in her placenta. Her SCN is an optical lattice clock; it is not a food clock. . It is a pretty remarkable gadget Nature built in our heads and our placenta.
It appears the simple addition of a neutron to H+ to the antenna system in the fetal thalmus is enought to screw up the connection of the thalmus to the heartbeat of the Earth. Without a proper connection to Earth, the fidelity of the music coming from the system fails. What does this FM station listen to? The Schumann resonace of the Earth. It is 7.83 Hz. That is the human alpha wave which is created in the thalamus. That is what causes all chronic disease in humans. It turns out adding neutrons to H+ is enough to do something to the fabric of space-time in the fetal brain that deforms the magnetic fields in the child’s ATPase to allow a child to emerge from a small pelvic outlet. This shows you how the smallest things make the biggest deal in you. Wild decentralized FACTS, that have been blocked in centralized science, I tell you. Sadly this science was shared with the corporations that were involved with the Industrial military complex in the 1970’s and 1980s.
THE PHYSICS WAS WORKED OUT FOR ME IN THE BASEMENT OF CHARITY HOSPITAL IN 1989.
The spacetime curvature for a charged static spherical body is given by the Reissner–Nordström metric. I’d suggest you carefully look at the next slide many many times. It shows you what Einstein’s special relativity really shows. This was critical for the development of technology to operate wirelessly using satellites to control them above Earth. This is why the GPA in you iphone actually works like your SCN does.
The picture above shows you time slows as one approachs the speed of light. The present moment your living right now reading this expands from a narrow sliver until it encompasses both the past and the future. This means that the idea of past and future is an optical illusion caused by the slowing of light by matter. It also means as light slows you get the idea there is a cause and effect on Earth but there is no real relationship. This is why BigHarma continues to use the idea that RCT is the gold standard for centralized science. The physics of special relativity tell you this is a bullshit story from the jump.
When you use these mathematics, you can feed in the value of whatever charge you want and calculate the time dilation as a function of distance from the charged body. If you do this you’ll discover something rather odd in Einstein’s field equations, namely, the charge reverses the effect of the mass in them.
This is why REDOX POTENTIAL is the key to chronic disease reversal and nothing else matters.
READ THAT LAST SENTENCE AGAIN.
This message was given to the Industrial military complex Big Pharma partners. They were told to add atoms to their drugs to limit the effect of charge. This helps the military use light to control people by using screen technology. It stole their dopamine reward tracts. I mentioned this pathway above. REVIEW IT.
That is what the target of MKULTRA was focused on after all the experiments where completed from 1953-1964. Those experiments continued until the early 1980’s by the CIA In Reagan’s admiistration. It was given to researchers who linked colleges via the ARPANET. This data was them shared by the Industrial military Microsoft’s Bill Gates and Apple’s Steve Jobs by DARPA in the 1980s. This was done by DARPA but pre- GOOGLE. . All that data was sitting in the basement of Charity Hospital where I did my residency. It all came from Tulane neurology and neurosugery experiments done for MKUTRA and by Sidney Gotlieb. I read them all before it was all destroyed by Huricane Katrina. The basement flooded and almost 160 years of medicine was lost in New Orleans.
How do cells increase their charge?
LIGHT DOES IT. Food has no effect on this.
Read it again.
LOOK MORE CAREFULLY AT THE SLIDE ABOVE AGAIN. It is why I posted it again. Do you remember how many times I have told you in podcasts that redox potential is formed by adding net negative charges in a cell? Melanin is the key way all mammals do it. Human biology relies heavily on this. Do you see now by examining the slide above closely how that charge in light frequencies can offset the bad isotopic variation in some of your tissues to make your clocks work better in the circuit that tells the brain what season it is? This is another universal law not subject to an RCT. BigHarma knows what your doctors do not. And they are not going to tell them either. It is encumbent upon you in getting this lesson and why it is being blocked from centralized MDs. It should explain to you why the Universities in ARPA founded HMOs and changes the way healthcare was delivered in the 1980s to put an Oracle between you and your doctor. That Oracle was a healthcare industrial complex player. The system was engineered to put a hospital CEO and an insurance executive decision tree between you and your doctor before ANYtreatment plan can be APPROVED by the governments agents in this plan. HMOs and the insurance industry were designed to limit the time a doctor can spend with you so you only have time to get a presciption that comes from BigHarma that changes the charge in you so you are more likely to follow the rules built by government. I will remind you that Medicare began post MKULTRA in 1968 under LBJ post JFKs assassination.
HOW WAS MKULTRA, LBJ, THE CIA, JFK, AND ISRAEL ALL LINKED TO THIS STORY?
I laid most of it out in the recent Danny Jones podcast but i saved the critical details part for my members so you can understand how it links to the creation of all modern disease epidemics.
THE MOST EXPLOSIVE SUMMARY
On July 30, 1965, President Lyndon B.Johnson signed into law legislation that established the Medicare and Medicaid programs. This created CMS for the industrial healthcare complex. It established Medicare, a health insurance program for the elderly who could and would be taken out, and Medicaid, a health insurance program for people with limited income to make sure they remained poor and sick to be controlled by the government for decades to come. LBJ signed this law in Indepence, Missouri. Why? This is where Manchurian Candidate Harry S. Truman was from.
This completed the medical tyranny portion of the New Deal Era that began under FDR and was linked to the the Manhattan Project story of how DARPA was created by General Groves. How long ago did the Industrial military complex decide to use medical tyranny as the attack vector of the US Constiution? 1911.
This was when the Flexnor report was launched to create BigHarma from Rockefeller’s break up of Standard Oil by Teddy Roosevelt. I had a lot to say about this in the Danny Jones podcast, but I will have a lot more to say about it in future podcasts once this information in that podcast is married in this summary and understood by the public.
Today, you are being highly preconditioned for compliance by the real findings that came from MKULTRA.
At the same time that CMS was created the ideas to taper the Ponzi scheme of money laundering from the public, the USA Executive Branch and the CIA helped pay off the MOSSAD for the JFK assassination by giving Isreal close to 300 pounds of uranium it needed to become a nuclear power. JFK, prior to his death, was going to send in the Atomic and nuclear regulators of the USA in to make sure Isreal never got an atomic weapon. This infuriated Israeli prime minister David Ben Gurion.
CAREFULLY EXAMINE THE PICTURE ABOVE AND NOTE THE DATE IT WAS TAKEN.
How did this story link JFK to Israel and the MOSSAD?
It began with an intelligence failure of the CIA and John Kerry’s father.
Documents published in Wilson Center from the National Archives now have shed light on a particularly notable intelligence failure: how Washington missed warning signs that the Israelis had a nuclear project underway, but also how the U.S. belatedly realized what the Israelis were doing, and how Eisenhower and his senior advisers reacted to this discovery. Among the new documents are:
The June 1959 Israel-Norway secret agreement providing for the sale of Norwegian heavy water to Israel (through the United Kingdom), transmitted by Oslo Embassy political officer Richard Kerry (father of former Secretary of State John Kerry).
Reports about information from a then-covert source — University of Michigan nuclear engineering professor Henry Gomberg — who learned that the Israelis had a secret nuclear reactor project that involved experiments with plutonium.
A telegram from the U.S. embassy in Tel Aviv reporting on Finance Ministry official Addy Cohen’s statement that “we’ve been misbehaving,” and one by an unidentified official close to Prime Minister David Ben-Gurion that the secrecy surrounding Dimona was unjustifiable and that it was “a stupid mistake on the part of Israel.”
Reports by U.S. Ambassador Ogden Reid on conversations with Ben-Gurion.
A State Department message to the embassy in Tel Aviv conveying irritation that the responses of the Israeli government showed a “lack of candor.”
Messages about a role for the International Atomic Energy Agency in inspecting and safeguarding Dimona. When Eisenhower left office Kennedy assumed this mess but he knew Ben Gurion for over a decade as the picture above shows. Now President Kennedy pressured the government of Prime Minister David Ben-Gurion to prevent a military nuclear program, particularly after stage-managed tours of the Dimona facility for U.S. government scientists in 1961 and 1962 raised suspicions within U.S. intelligence that Israel might be concealing its underlying nuclear aims. This went on during the Cuban Missle crisis and Kennedy was worried about nuclear proliferation post WW2. Kennedy’s long-run objective, documents now show, was to broaden and institutionalize inspections of Dimona by the International Atomic Energy Agency.
Of all U.S. leaders in the nuclear age, John F. Kennedy was the nonproliferation president. Nuclear proliferation was his “private nightmare,” as Glenn Seaborg, his Atomic Energy Commission chairman, noted many times in interviews. Seaborg discovered Plutonium for the Industrial military complex leaders.
On 30 May 1961, Kennedy met Ben-Gurion in Manhattan to discuss the bilateral relationship and Middle East issues. However, a central (and indeed the first) issue in their meeting was the Israeli nuclear program, about which President Kennedy was most concerned. According to a draft record of their discussion in the National Archives, which has never been cited, but has now been released for the first time by the Wilson Center, Ben-Gurion spoke “rapidly and in a low voice” and “some words were missed.” He emphasized the peaceful, economic development-oriented nature of the Israeli nuclear project.
Nevertheless the note taker, Assistant Secretary of State Philips Talbot, believed that he heard Ben-Gurion mention a “pilot” plant to process plutonium for “atomic power” and also say that “there is no intention to develop weapons capacity now.” This information stunned JFK. Ben-Gurion tacitly acknowledged that the Dimona reactor had a military potential. The final U.S. version of the memcon retained the sentence about plutonium but did not include the language about a “pilot” plant and “weapons capacity.” Anyone who understands basic nuclear physics (see my Patreon Thorium blog on BTC) knows the only reason to use plutonium as a fuel is because you can make military grade uranium for nuclear weapons.
The differences between the two versions suggest the difficulty of preparing accurate records of meetings. But whatever Ben-Gurion actually said, President Kennedy was never wholly satisfied with the insistence that Dimona was strictly a peaceful project. This really was the main precursor to his death in my estimation. Neither were U.S. intelligence professionals. A recently declassified National Intelligence Estimate on Israel prepared several months after the meeting concluded that “Israel may have decided to undertake a nuclear weapons program. That CIA information was leaked to the mob in 1963 after the Bay of Pigs invasion went south. This information was critical to Santo Trafacante because it allowed him ot gain leverage over Castro and the attempts of the Industrial military complex to kill him versus JFK. Trafacante ensured it was JFK that would pay the ultimate price. He viewed his appointment of RFK Sr as Attorney General as the ultimate act of betrayal to the mob bosses in Chicago, New York, Florida, and New Orleans. In fact, Carlos Marcello was on trial as JFK was shot in Dallas on November 22, 1963 because of an RFK Sr prosecution that Joe Kennedy swore would never happen if the mob swung the swing state of Illinois to JFK in 1960. These details matter big time to understand how things really unfolded.
ISRAEL had massive amounts of agents in the USA in the Jewish mob before Israel was even made a country in 1948. Prior to being a country they had a massive intelligence capability world wide that linked banking, science, and gangsters who were all Jewish.
This was the main reason the Jewish gangsters were eventually made MOSSAD agents after 1948. They fed information to Israel that the Chicago and NY families got on Joe Kennedy and his sons. As a result, Meyer Lansky, Jack Ruby Rubenstein, Carlos Marcello, and Santo Trafacante became involved with the assassination of JFK , along with Nixon, the Bush and Hunt Oil families and Clint Murchison linked to Groves Industrial military complex. Texas was also a swing state and the Oilmen were pissed that Kennedy was catering to Blacks in civil rights and was soft on the Communists in Russia and Cuba.
Santo Trafacante was among the most powerful Mafia bosses in the United States when JFK was a Senator in the 1950s. He headed the Trafficante crime family from 1954 to 1987 and controlled organized criminal operations in Florida and Cuba. He was also a MOSSAD asset for reasons I laid out in the Danny Jones Podcast. He was a double agent for the CIA and MOSSAD in the 1960s due to drug laundering to replace what the Chicago and Florida families lost in the casinos due to the Cuban Revolution of 1959.
Castro enlisted Santo to pay back the CIA for trying to kill him 9 times by joining forces with Traficante to turn Americans into drug zombies. Trafacante was the key link the Italian mob to the Jewish in the USA. This is how the MOSSAD got the information that JFK life was at risk. He also was in charge of monitoring the behavior of Judyth Vary Baker under Dr. Alton Ochsner care on Magazine St in New Orleans. He also linked the Chicago mob the NYC mob of Bonanno crime family. This family is known as the American mafia. They are constructed of 5 families all based in New York. These Families had connections with FDR Jr son as a Senator from NY.
Carlos Marcello was a gangster who was based in New Orleans but tightly linked with Sam Giancana of Chicago. Both worked with Trafacante in Florida and Chicago. He knew Lee Harvey Oswald since Oswald grew up in New Orleans. In 2022 more government documents were released which showed us how US law enforcement was linked to the Italian mob. At least nine files released refer to Chicago circa 1963 and several individuals linked to the Outfit.
Most notably among them are former Chicago police officer and Cook County Deputy Sheriff Richard Cain (see above). Cain had deep connections to Chicago organized crime according to investigators, especially ruthless mob boss Sam “Momo” Giancana. Joe Kennedy, father or JFK, asked for Sam’s help in getting Kennedy elected in 1960 through his connections with labor unions linked to Jimmy Hoffa. Illinois was a He was promised under JFK presidency there would be a laissez faire action plan towards organized cirme. This is why Sam helped Kennedy’s father. It is clear today that JFK or RFK was not given this information. It is also clear that RFK Jr knows this information today.
In 1965, the MOSSAD was paid off when Isreal got uranium from LBJ and CIA as payment for their assistance. Did Israel steal bomb-grade uranium from the United States or were they given it by the USA?
After a 1965 inventory, NUMEC was found to be missing about 100 kilograms of bomb-grade uranium, even after accounting for all processing losses. The close personal and commercial ties to Israel of the plant owners and operators raised suspicions that remained unresolved. The affair of the missing bomb-grade uranium was revived in 1976. The newly formed NRC in 1976 UNDER fellow Warren Commission member Gerald Ford who replaced Nixon in 1974.
The NRC was in the process of writing licensing regulations for commercial fuel firms—of which NUMEC was one—and had heard rumors of possible theft in the 1960s from NUMEC’s Apollo facility. NUMEC = Nuclear Materials and Equipment Corporation. NUMEC is based in Pennsylvania. There is a reason this plant was targeted by LBJ, CIA, and Israel. The partners in the JFK crime syndicate were in control of this location. Arlen Specter the lawyer who came up with the one bullet theory in the Warren Commission was given the Senator postion by the Industrial military complex by using election stealing capabilities in the 1960s. The same ones were used to give the election of JFK over Nixon in Illinois and Texas in the 1960 election.
The NRC asked for a CIA briefing. This was by design because President Ford knew the CIA was involved in the theft to pay Israel off. Duckett startled the NRC group with CIA’s conclusion that the missing uranium was in Israeli bombs. The NRC chairman informed the White House, and President Ford took an iimmediate nterest in the case because Duckett was not supposed to tell the truth. He was not involved in the JFK assassination.
Ford’s Attorney General, Edward Levi, discovered that the Atomic Energy Commission (AEC), the NRC predecessor nuclear licensing agency, had previously convinced the FBI not to open a criminal investigation into the material’s disappearance. Why would he do this? Levi was the first Jewish Attorney General of the United States. He was also a MOSSAD asset.
Levi was a Yale Graduate with ties to Israel. In fact, he was a Sterling Fellow at Yale and that dates his affliation with Israel all the way back to 1864 and B’nai Birth. B’nai Birth is the previous group now called AIPAC. Before AIPAC they were the ADL. What is the history of B’nai Birth? This group was loyal to the Royal Family before the Civil War in the USA. As a Yale graduate he was linked to the Bush oil family who was an early partner of the Industrial military complex General Groves put together from the ONI and OSS before the CIA was formed. Preston Bush, the father of George H Bush, was a banker who worked Brown Harriman in NYC who worked with the families who controlled money in the US Federal Reserve. This was established in 1911.
During Levi’s term as Attorney General, he issued a set of guidelines in 1976 to limit the activities of the FBI. This was done in repsonse to Nixon being forced to resign over wiretapping during Watergate. The CIA was furious that the FBI turned on Nixon and this was one way the DOJ in the executive branch was used via lawfare. These Levi guidelines required the FBI to show evidence of a crime before using secret police techniques like wiretaps or entering someone’s home without warning.
How connected to the criminal cabal was Levi? His guidelines were QUICKLY replaced by new ones issued in 1983 by Ronald Reagan’s Attorney General, William French Smith. Levi urged President Ford to appoint Robert Bork, who was his former student and Solicitor General to the SCOTUS. The real reason Bork was not elevated to the SCOTUS is because many democrats knew that Levi and Bork were linked to the conspiracy that assassinated JFK. That did not stop Levi. He told Ford if Bork is blocked he should nominate fellow Chicagoan John Paul Stevens to the United States Supreme Court, and Ford followed his advice. Did you know that Levi later testified in support of Bork at his confirmation hearing for the SCOTUS? Did you know who else was linked to Levi and Israel?
Serving under Levi at various times in his career, in various high staff positions, were such people as Rudolph Giuliani, Robert Bork, Antonin Scalia, Rex E. Lee, and Arthur Raymond Randolph. That is how close the ties with Israel and the MOSSAD are to the USA. It is also why I told Danny Jones why Donald Trump made the JFK achives public. As president, he found how his friends in banking, Wall Street, and real estate were to the JFK assassination attempt. They are the people behind the warning shots being made right now to DJT. I believe if they wanted him dead, he’d already be dead. I think they think they can control Trump if they make it clear to him during this election cycle that he needs to follow the playbook Levi built out in the 1970s. Joinign forces with RFK Jr is why they went after him at Butler in Penssylvania. It should not surpise anyone of you now why it happened in Pennsylvania. The industrial military control over this state and it politicians is close to air tight.
HOW TIGHT IS PENNSYLVANIA in this story?
I told you that NUMEC is in Pennsylavania. Did you know that Zalman Shapiro the owner of NUMEC attorney was Arlen Specter, the Senator for Pennsynvania? Amazing coincidence or was the something more important to this story the USA government wanted buried forever?
So the chain of evidence went from Ford to Bush to Carter to the CIA or did it?
Carter instructed his national security advisor, Zbigniew Brzezinski, the father of modern day broadcaster Mike on MSNBC, to deal with the NUMEC matter in the context of the impending public release of MUF data. Brzezinski’s staffers John Marcum and Jessica Tuchman posed questions to the CIA about the NUMEC affair. This is when key data magically went missing to President Carter.
WHAT WENT MISSING?
Carter was never told about the links of Israel to NUMEC.
Brzezinski shared the information with 2 staffers but not Carter. The two staffers told Brzezinski that environmental samples taken by the CIA in Israel in 1968 contained highly enriched uranium, whose enrichment level was so high it pointed to the Portsmouth, Ohio, uranium enrichment plant as the source. Portsmouth was where NUMEC obtained uranium stock for its naval fuel products. Why he never told Carter is likely tied to his ideology as Polish and hating Russia and this marries to General Groves ideology linked to Stalin.
During the Apollo affari of 1965 the president of NUMEC, Zalman Shapiro, had frequent contacts with Israeli officials, including a science attaché “thought to be an intelligence officer,” and received unexplained VIP treatment in Israel. None of this information went from Brzezinski to Carter.
In fact, Shapiro was by then known to have had contacts with Israel’s head of military intelligence and the head of its nuclear weapons program. He later acknowledged knowing Binyamin Blumberg, head of Israel’s “bureau of scientific liaison,” which engaged in high-risk intelligence capers. On one strange occasion in 1968, Shapiro hosted an Israeli intelligence foursome at the Apollo plant. One was the Mossad agent who headed the team that spirited former Nazi leader Adolf Eichmann out of Argentina and who later ran Jonathan Pollard’s spying on the United States for Israel.
For those of you who do not know, the Mossad is the Israeli agency that handles foreign intelligence collection and covert action. It is Israel version of the CIA. Another was that agent’s deputy in the Eichmann kidnap, who went on to become head of Shin Bet, Israel’s internal security service. A third was Mossad’s director of technical services. The last was Israel’s science attaché, who had held a senior position in Israel’s nuclear weapons program. More amazing coincidences in this story do not you think?
How did I know all this history and link it as a young man before I became a doctor in New Orleans? The man who i have spoken about in many podcasts who taught me everything I know about fiat banking and fractional reserve banking was named Theodore Sterling of the Sterling National Bank on 57th Street and Madison Ave as I grew up in NYC. He told me this history when he told me how banking really operates in the USA. Read the hyperlink on him you just passed before going further.
What happens when Ford leaves office? How did the story remains toghtley controlled by the industrial military complex?
Everyone involved was linked to the Warren Commission.
After the 1976 election, the Ford White House alerted the incoming Carter administration to the NUMEC affair. In December 1976, according to a July 1977 National Security Council memorandum, then-CIA Director George H.W. Bush briefed President-elect Carter on the case. Congress had been pressing for public disclosure of records of large unexplained losses of bomb-grade material (in government parlance, “material unaccounted for,”or MUF) from the government’s nuclear weapons complex. The Carter White House feared the story was sure to hit the headlines if there were any suggestion of Israeli theft from the NUMEC facility.
And any disclosures about Israel’s bomb program would of course have threatened the Carter administration’s Middle East policies between Israel and Egypt. If you really want to know the truth, I have always believed Israel agreed to this peace deal reluctantly to bury the Apollo affair once and for all. But that affair is the gift that keeps giving. It’s secrecy is linked to the real story of how the MOSSAD was involved in 9/11 Tower knockdown fiasco during the George W. Bush presidency. See how the story all begins to make sense when you really see it laid out as it happened. Today’s history books will never let you see how it is all linked.
MOST OF YOU HAVE NO IDEA HOW NEFARIOUS OR HOW OLD THIS PLAN IS IN THE HISTORY OF THE USA.
You’re going to be retooled with knowledge and reschooled with Nature’s wisdom today. Share this decentralized wisdom with the world.
Do we need to know our “WHY“?
I think we do.
Without knowing our why I believe the attainment of Optimal will not happen.
I was getting ready for a medical talk earlier this year and I was jotting some things down about how my surgical judgement has changed from the time I left residency to the present and going over my practice patterns for the talk. It dawned on me the practice of medicine is very much like the practice we use in life to become a more imporved version of ourself. Inevitably, practice isn’t the thing you do once you’re good. It’s the thing you do that makes you good. I was surprised about the crowd of people I was asked to speak to. The mayor was in the audience and so was some the state health department officials. I had no idea who i was speaking too. I thought it was going to be mostly patients and PCP’s in my new referal base for my new job. When I saw that my audience was quite different than I anticipated I scrapped my “neurosurgery talk plan” on minimally invasive spine surgery and decided to talk about how my “what”, neurosurgery, led me into my “why”, my real passion………Quantum biology.
The response I got from yesterday’s talk even astounded me. I think I realized something about myself yesterday. When I speak from the heart I connect people with nature. That connection is the key to getting my flock well. This is the basis of becoming a mitochondriac. I realized this week that I help people find their “why” in healthcare via biophysics so their wellness manifests. Do you need some clarity on how to move your “what’s”, to the key “why’s” that will eventually move your decision making process to focus on only the things that matter in your life to keep you well? Then reach out to me and join my flock of mitochondriacs right now.
SO GIVE ME SOME EXAMPLES DOC
“The antagonistic pleiotropy (AP) theory posits that aging occurs because alleles that are detrimental in older organisms are beneficial to growth early in life and thus are maintained in populations.
Although genes of the insulin signaling pathway likely participate in AP, the insulin‐regulated cellular correlates of AP have not been identified.
Answer: young humans before full myelination in brain needs deuterium to complete growth but once myelination is complete then mitochondria need to deuterium deplete to control growth and metabolism pathways into adult life. This is the control system for the insulin signaling systems built into the placenta. The proxy for the system is salinity measures. The more salinity in the system the more UV light can be captured in the system to drive morphogenesis.
As deuterium fractions in tissues rise as we age, salinity measures lower. This means we need more sunlight to offset the risks of aging. This is why those with insulin resistance have high BUN/creat ratios and why they are more apt to have SIADH, HTN.
Sodium transporters brings Vitamin C into the brain for TCA cycle protons recycling to lower the KIE of deuterium in TCA intermediates as we age to control volume changes in the matrix. This is why SIADH is associated with cognitive decline in TBI and aging.
MORE DECENTRALIZED LESSONS?
Na also brings calcium into bone in cases of osteopenia. Low Na = low Ca and Mg = mitochondrial dysfunction. Low salinity in CSF and the body is a feature of all nnEMF exposure in environments.
The higher nnEMF in your environment the lower salinity becomes in blood and CSF and lower salinity = sleep disorders. Most people with high insulin levels due to artificial light exposure also are afflicted with sleep apnea due to deuterium rise cause by the altered Vitamin A signaling. The slide below makes this case.
BUN rises as salinity decreases because deuterium fractions are increasing. Salt from our eccrine sweat glands helps cool our body because it augments skin vasodilation ( like UVA light does).
As man made blue light exposure of skin rises, dopamine creation in the eye and brain drop and our salinity in CSF drops and obesity manifests easier due to deuterium content in the choroid of the eye and the skin and subcutaneous fat mass due to melanopsin.
The TRP receptors of melanopsin are affected negatively by deuteration by altered salinity.
What are TRP receptors?
Transient receptor potential channels, were first discovered in 1969. They are multimodal ion channels that act as sensors of photic, chemically, or physical toxic stimuli. These channels are widely distributed in various tissues and play a variety of roles. Centralized science is still yet to realize how these channels work with light. Blu elight varies with the time of the day and this system measures these changes accurately to tell time at the quantum scale.
It all makes sense when you understand bio-physics of the kinetic isotope effect (KIE) of deuterium in the mammalian system. Centralized MDs have no clue about this work. Most have no clue why PUFA’s in fake fats are really bad: they increase proton leakage in mitochondrial matrix and allows more deuterium in matrix than should be there and this raised heteroplasmy rates to cause disease.
SUMMARY
How do we link the lessons I have taught you over the years to light?
CLOCK and BMAL1 are positive regulators of circadian gene expression in humans, and PER and CRY are the negative regulators that operate under day and night cycles.
Blue light and nnEMF liberate Vitamin A and when it is free in the system it sets off a chain of events. None fo them are good for the non visual photo receptors. For examples, it becomes an aldehyde that destroys the small molecule modulators of the mammalian circadian mechanism shown in the picture above. Once the molecular clock goes awry in a tissue with poor light signaling the implications for many neolithic diseases spiral out of control. What are some of the Vitamin A proteins involved in this downward spiral?
They are called retinoic acid receptor-related orphan nuclear receptors or RORs for short. The RORs have several isoforms too called RORα-γ. These proteins are also under the transcriptional control of CLOCK/BMAL1 heterodimers. This is bad news for the circadian control of PER.
SIRT1 regulates the activity of BMAL1 and CLOCK, two circadian transcription factors, which target NAMPT, an enzyme that synthesizes NAD+ in cytochrome 1. And in a curious feed-forward mechanism, CLOCK and BMAL1 enhance SIRT1 expression… genetic deletion of any of these players induces insulin resistance in humans. This is how chronic blue light exposure causes diabetes. Food does not cause it. LIGHT DOES. Food exacerbates the situation only because carbohydrates make 50% less water at CCO compared to fats.
Sunlight is a NATURAL calcium channel blocker that alters the firing rate of voltage-gated channels on cells. SUNLIGHT in the AM increases the amount of PER protein that is made in the cytoplasm to get to a critical concentration where it enters the nucleus. This alters the free radical signal in mitochondria. Recall all free radicals have one unpaired electron and this makes them magnetochemicals that contol timing decisions in the cell. They act as a logic gate to chose the proper metabolic pathway to use.
This is done in a few ways with sunlight. AM sunlight increases PER, and later in the AM UVA light shows up = nitric oxide (NO) release = lowers BP and controls stem cell depots and the blue light in sun balanced by red light also stimulates melanopsin relaxation of the arterioles in our skin as another collateral effect. This one reason why just going outside in the sun and clean air reduces depression and anxiety because sunlight slows breathing and improves oxygenation. When PER is not made by sunlight depression and suicide are more predictible. Indoor living and ALAN lead to depression and suicide because PER is not made by AM sunlight. the picture shows you the effect in the SCN. It is devasting to the signaling in the leptin melanocortin semiconductive pathway. Smash that blue hyperlink now.
Breathing indoors requires us to use suboptimal air which is not moving or mixing with the wind or interacting with the sun so its charge is lowered. This poor oxygen level = pseudohypoxia = poor PER creation in the AM —> NAD+ drops = higher heteroplasmy = you age faster = more sympathetic activation = more PVN firing = less vagal tone = you get sick quicker indoors. More than 90% of you live inside.
As a result, SIRT 1 lowers with INDOOR living = alters PER cycling in the SCN and every tissue from the cytosol day and night from the nucleus.
Why is this a big deal? When PER one cycle is off so is NAD+ at cytochrome 1 and this causes advanced aging in man due to heteroplasmy because deuterium is let into the mitchondrial matrix where it is not supposed to be!!!
NAD+ is one of the more immediate players in cytochrome 1 that is a huge driver of circadian biology in humans. It is the coenzyme called nicotinamide adenine dinucleotide (NAD+). It participates in a variety of redox reactions in the matrix that help generate DDW. Solar exposure and fasting work with light frequencies to slow ECT flow and this can increase the intracellular NAD/NADH ratio if the light environment is dominated by sunlight. It won’t do this with artificial light. It lowers NAD+. This is what sets off a cascade of circadian events that can destroy tissues because they involve epigenetics and the regulation of growth and metabolism of man. LIGHT DOES THIS. NOT FOOD OR FUELS.
SUN + fasting -> PER cycling is controlled by AM sunlight —> raised NAD+ -> SIRT1 -> BMAL1/CLOCK -> NAMPT -> NAD+ is increased.
You’ve now been retooled with knowledge to be reschooled with wisdom. Share this decentralized wisdom with the world.
I’ve received a ton of DMs and emails about my discussion with Dr. Alexis Cowan on our second podcast on T cells and autoimmunity, so I decided to answer them here.
How do I know that the external process on the integument controls the biochemistry process on the interior? Do you know that supplementation of Vitamin D does not work the same in those with heavily melanated exteriors compared to those with Fitzpatrick one and two skin?
According to the article above, “Doctors frequently prescribe supplemental vitamin D,” Freedman said. “However, we do not know all its effects and how they may differ between the races. The bottom line is that racial differences in calcium handling are seen, and black and white patients have differing risks for bone and heart disease. We should more clearly determine the effects of supplementing vitamin D in black patients with low levels based on existing criteria. We should not assume that the effects of supplementation will be the same between the races.” Again, this is not a racial story; it is a story of the biophysics of melanin and how POMC is only translated by UV light frequencies.
ARE THERE ANY OTHER LINES OF EVOLUTIONARY EVIDENCE THAT THESE THINGS ARE LINKED TO OUR INTEGUMENT?
YES THERE IS IN DAIRY TOLERANCE. This is food loaded with calcium. HYPERLINK
This explains this fact: 80 percent of all African Americans and Native Americans are lactose intolerant. They have more melanin in their integument. Over 90 percent of Asian Americans are lactose intolerant, and it is least common among Americans with a Northern European heritage who have Fitzpatrick one or two skin.
This important distinction must be made in El Salvador, where 95% of the population has melanated skin. This implies that supplemental Vitamin D is not a wise choice for them.
Brown & Black races generally have lower vitamin D levels than whites, partly because their darker skin pigmentation limits the amount of the vitamin produced by sunlight. This is protective in environments where melanated skin is optimized.
IMPLICATIONS OF THE SURFACE BIOPHYSICS?
Despite these lower vitamin D levels and dietary calcium ingestion, browns & blacks naturally experience lower rates of osteoporosis and have far less calcium in their arteries based on published studies. Studies further reveal that brown/black patients with diabetes have half the rate of heart attack as whites. This shows us that melanated skin may be a protective effect from blue light and nnEMF exposure. This also shows us that lower levels of calcified atherosclerotic plaque in brown & blacks are associated with a lower risk of heart disease. The irony is in the stroke and heart attack belt in the US, browns & blacks in the general community have higher rates of heart attack than whites. This implies that their circadian mismatch to a higher latitude likely explains their decreased longevity. This is why my pinned tweet exists. Vitamin D is not the best end-all story in longevity. It can be used as a good proxy for whites, but we should not assume the same for browns & blacks. Browns & blacks clearly get disease faster and more severe than whites at the same latitude. This again points to a lack of proper sunlight being the main culprit in disease creation.
Humans have CYP11A1-derived secosteroids in their epidermis and serum. What does it do?
CYP11A1 drives a novel magnetochemical pathway in the vitamin D3 (D3) and 7-dehydrocholesterol (7DHC) metabolic pathways in humans. CYP11A1 initiates it. The CYP11A1 SNP has been previously characterized in vitro papers. It has also been proven to occur in vivo in humans. The researchers in those papers analyzed samples of human serum, epidermis, and pig adrenals for the presence of intermediates and products of these pathways. What did they find, and why did it sway me that the electric state of our membranes is giving direct feedback control to metabolism below inside of cells?
In the paper linked below, the epidermal, serum and adrenal samples showed the presence of D3 hydroxy-derivatives corresponding to 20(OH)D3, 22(OH)D3, 25(OH)D3, 1,25(OH)2D3, 20,22(OH)2D3, 20,23(OH)2D3, 20,24(OH)2D3, 20,25(OH)2D3, 20,26(OH)2D3, 1,20,23(OH)3D3 and 17,20,23(OH)3D3, plus 1,20(OH)2D3 which was detectable only in the epidermis.
Serum concentrations of 20(OH)D3 and 22(OH)D3 were only 30- and 15-fold lower than 25(OH)D3, respectively, and at levels above those required for biological activity as measured in vitro.
They also detected 1,20,24(OH)3D3, 1,20,25(OH)3D3 and 1,20,26(OH)3D3 in the adrenals. Products of CYP11A1 action on 7DHC, namely 22(OH)7DHC, 20,22(OH)27DHC, and 7-dehydropregnenolone, were also detected in serum, epidermis, and the adrenal gland. Thus, research has shown that humans have novel CYP11A1-derived secosteroids in the skin, serum, and adrenal gland. This SNP likely has much to do with why we deleted the Vitamin C gene in the human genome. Moreover, as I told Dr. Cowan, based on the concentrations found in the papers, their biological activity suggests that they act as hormones in vivo.
T cells undergo positive and negative selection in the thymus through T cell receptor (TCR) recognition of peptides presented on major histocompatibility (MHC) proteins. Although the genes encoding MHC proteins are among the most polymorphic, most vertebrates express relatively few MHC alleles individually. Genes only act to alter metabolic flux, as the picture above shows. How the number of MHC alleles expressed affects thymic selection and the TCR repertoire on an individual level remains unclear to centralized science. Decentralized science has a different viewpoint. I think sunlight on the skin changes our microbiome and sculpts it to create free radicals and light changes from the microbiomes, creating a variability response in our CD4 and CD 8 cells.
SUMMARY
This is my current hypothesis. I believe sunlight creates T-cell variability in our skin and gut surfaces, as I mentioned below with Dr. Cowan. I think sunlight stimulates MHC heterozygosity, which improves immunity by reducing receptor variability in T cells. MHC heterozygosity limits T cell receptor variability in CD4 T cells. T cells are essential for recognizing infectious agents through their receptors (TCRs), which engage peptides derived from invaders or auto-antigens bound to major histocompatibility complex proteins (MHC). While much research has focused on MHC peptide specificity, the impact of MHC on the expressed TCR repertoire, especially in MHC heterozygotes, has not been well studied in centralized science. Understanding this decentralized link in medicine should guide personalized autoimmune monitoring and better predictive responses to various new treatments. Sunlight exposure to 380nm and 1280 nm light is mandatory in any autoimmune reversal. It should be from sunlight and not any manmade light.
The blog’s parting thought is about pharmaceuticals for autoimmune conditions: The human condition does not require that you take drugs that pharmaceutical companies sell. You do not have a pharmaceutical deficiency or a “chemical imbalance.” BigHarma sells these ideas to keep you believing their BS. Pharmaceutical companies demand that you consume what they sell. You can live strong and healthy without them. Learn how and do it. Decentralized medicine and healthcare teach these techniques.
Time is your most valuable asset in life and links to your choices in health span and how you use your monetary energy. Why doesn’t the “food guru” get this concept? They spend too much time under ALAN while in a gym reading flawed nutrition data and not enough time reading about the physics of organisms. The erasure of information drives entropy in cells, and this process increases over time. This is a consequence of having a ratio in our universe of 1 billion light photons to every atom. This is why LIGHT> food. All molecular clocks that control circadian biology are flow meters for this entropy measurement. This should tell the ripe mind that circadian biology’s light and dark cycles are the key to the biological ENIGMA CODE of all modern diseases, especially autoimmune conditions.
Be wise and get a job so your life’s art can be free or at least close to it. Do you. Create for yourself. Do not create for the audience. In fact, I will tell you that if you do act like the customer is always right, your life’s work will never resonate to every inch of this planet. So many creators have been miserable or given up because they were taught your creativity has to pay for your life, or you’re no good. That is an idea that had to be born in a food guru’s mind. It is devoid of wisdom and impotent.
Centralized Medicine as it practiced today is an applied science. It is not foundational science. If that offends you, good. You’re ready for this lesson. The centralized experts on act like it is foundational to Nature. Nothing can be further from the truth. Glymour and Stalker argued at the turn of the 21st century that “medicine in industrialized countries is scientific medicine.” I remember reading it and thinking this has to be the most ignorant statement I have read post training. I read it 5 years before the insights came to be in my Quilt document. It might have been my fuel to burn this idea to the ground.
Although centralized physicians’ medical practice is not itself science, it is based on a manufactured science and on training that is supposed to teach physicians to apply scientific knowledge to people in a rational way. Unfortunately that mode of dissemination has been polluted by the Oracle problem at the core of centralized medicine, namely, BigHarma and its curriculums. This distinction between understanding nature and power over nature, between pure and applied science, was first made by Francis Bacon in his Novum Organum of 1620. It appears to me centralized science has been going off the rails ever since.
Thomas Huxley pointed out in his address at the opening of Mason’s College in Birmingham, England, in 1880 that applied science is nothing but the application of pure science to particular classes of problems. No one can safely make these deductions unless he or she has a firm grasp of the principles that rule science. No one in centralized science does. Yet, the idea of the practice of clinical medicine as an art persists. What is this?
Does it amount to anything more than romantic rhetoric—a nod in the direction of modern transhumanitarianism? I think that is exactly the problem.
I was taught basic medicine using Cecil’s Textbook of Medicine.
There is a discourse on medicine as an art at the start of Cecil’s Textbook of Medicine. It focuses on the patient, a fellow human seeking help with a health-related problem. This leads to the observation that, in order to be considered medicine as an art, the human faculty must be its primary and characteristic instrument. What are the aspects of faculty that matter?
The physician, in fact, is invited to be part of the treatment process in listening, empathizing, informing, and maintaining solidarity. We wouldn’t argue with the usefulness of these properties, but they also describe moral dimensions of care-we listen because we respect people-and skills. As with technical skills, interpersonal skills can be lacking frequently. Like any practical technical skill, they can be observed, taught, tested, and their value assessed.
Medical teachers probably say much the same thing about their third mantra: attitudes. While these may be more dependent on physicians’ upbringing and personalities, attitudes can be changed with education or appropriate legislation, can be observed and scored, and can be evaluated in their contribution to patient care or diagnostic technique—at least in principle and even if these are crudely done. Part of the art of clinical medicine may lie in these areas, but not exclusively so: the art is not just practical performance. I want to suggest to you right now that the art and science of medicine are inseparable, part of a common culture. I just did a talk at the Palestra Society in El Salvador about this idea. Decentralized science requires personal participation in knowledge; knowing is an art.
In the accumulation of such knowledge, centralized physicians act just like engineers; they share experiences individually through meetings and publications. Within the community of its discipline, this intersubjectivity establishes the objectivity of science: it is knowledge that can be publicly tested using a PEER review system. Today, that system is flawed by the Oracle problem of journal editors who decide what is fit to publish and what should perish. It is how centralized science censors its best minds who have the best ideas. This approach can be summed up as a doctrine of standard empiricism in which the specific aim of inquiry is to produce objective knowledge and truth—and to provide explanations and understanding. Science as pure science is knowledge of the natural environment for its own sake, or rather, for understanding. Science as applied science or technology is the exercise of a working control over it. Such is the state of centralized medicine. The collateral effects of this are massive and the implications are larger for the public health
Centralized medicine currently believes its methods, scientific thinking should—must—be insulated from all kinds of psychologic, sociologic, economic, political, moral, and ideologic factors that tend to influence thought in life and society. Without those proscriptions, objective knowledge of truth degenerates into prejudice and ideology. And the COVID EPOCH proved that.
Centralized scientists & physicians like to bask in the reflected glory of medicine as a scientific undertaking that transcends national barriers. The industrial healthcare complex, WHO and the industrial military complex that powers it all supports a vast number of international academic meetings & international specialist societies who set down the guidelines to practice by. They are rigid and support the paradigm in power making the rule while the public health falls into the abyss of chronic disease on every continent.
DECENTRALIZING SCIENCE
Life is like a photograph; it often develops from the negative in life. I got this insight from my observations in museums as a child. It made me realize the truth is best found in our failures. So I decided to jump head first without wings into centralized medicine’s biggest failures.
I realized the human mind is a thinking machine. I then thought about its failure as our weakest link in centralized medicine. I thought to myself, “can machines think better than us?”
In 1950, Alan Turing had an answer to that question—a computer was capable of “thought” if its output was so convincing that a person interacting with it couldn’t distinguish its answers from those of a real human. The concept, known as the Turing test, has regained new prominence today as some argue that this new generation of Artificial Intelligence does in fact pass the Turing test. That is when I realized that transgenerational epigenetics, our offspring, is the HUMAN version of the Turing test.
Turing had a unique and brilliant mind. He apparently had the same thought as I did in 2005. His idea came to him in 1930.
At boarding school, a teenage Turing formed a close bond with his classmate, Christopher Morcom, over their love of science. When Morcom died unexpectedly in 1930, Turing was devastated. In letters to Morcom’s mother, he wondered whether Morcom’s mind could possibly live on without his body.
The loss changed Turing, plunging him into a philosophical, mathematical, and scientific investigation of the human mind that would last a lifetime. My day at the foot of this statue did the same thing to me. That is the day I found my way home. Back to Nature. I realized everything was decentralized in us.
I realized I had to come off my throne, and had to leave the body of my education alone. I knew I had to change. And I did.
WHAT HAPPENED ON THAT DELTA FLIGHT?
I figured out where all the modern disease epidemics began rapidly.
Most of you know by now, I am hell bent on changing the process of how centralized medicine is practiced and healthcare delivered. Right now my sphere of influence is small and I can affect my patients and family with my decentralized thoughts. Thinking about how to do this is a messy process when the end point is not well defined. I do know that it must change, but for now I am going to trust the processes Nature exhibits and transfer my knowledge to people as I move through life. Some paradigms change slowly and others happen like a Tsunami. I have decided to embrace change and try to become the change I want to see in medicine now. I realize I cannot control it, but I know I can work in harmony with it and even cultivate the vision I see for it. I used to worry about healthcare reform and how it might end up after Washington DC tried to control it, but I realized that if I could not control it neither can they. I found some solace in that.
It has allowed me to focus on what really matters, and that is decentralized health, quantum biology, and the physics that underpins evolutionary medicine to mold the future of healthcare. Patients want results and are now mandating they happens on their terms. This cannot happen. Patients need to realize how their light environment hijacks their ability to think. Purple and red light from the sun are critical in this dance, and modern lighting has subtracted both of these colors from the rainbow of modern lighting. I love this change in thought because it is fundamentally decentralized at its core. Light and dark environments control our biology. It should never have been usurped by the ideas the profiteers of BigPharma. BigPharma institutionalized their ideas like a gang member does to neighborhood. The BigPharma gang’s neighborhood is a medical school curriculum and its targets are centralized phyicians and all the patients they treat.
Centralized healthcare is simply gang warfare at scale. Moreover, it became government supported during the Vietnam War era by LBJ and NIxon and was codified in the dietary guideline of America. Politicians in Washington DC never wanted food inflation to ever be an election issue again. This is when we embrace cheap manufactured food to the American popualtion at scale. This occured at the same time we debased money, removed the gold standard and allowed the Nixon Shock to have its full effect on centralized medicine and created a powerful industial healthcare complex built to take advantage of sick care using the Cantillon effect. This endless siege of terror in healthcare continues to be funded almost exclusively through money printing and BigPharma continues to sit very close to that printer. This eefect can be seen by policy makers in Washington DC who have looked to centralized medicine further since the Food Pyramid was built by the architects of centralized destruction.
Eliminating cheap money from this system will eliminate the reliance on manufactured cheap foods. Decentralized ideas on health and money ultimately brings peace in the war on the public’s health because it stops incentivizing the problem at the core of the industrial healthcare complex. It also uncouples the link of government to science that began in the Manhattan Project under the control of General Groves. This is when the industrial military complex got the idea that marrying BigScience to industrial military contractors was a good business idea for the military. In his farewell speech to the Union, President and General Dwight D. Eisenhower tried to put the brakes on what General Groves began but was unsuccessful. He warned the American public of the coming collateral damage of such a decision but we did not heed the warning. Today, the chronic disease peidemic is a result of that mistake.
Physicians and patients who embrace decentralization in mediicne, and know that modern medicine is more centralized. The reliance on technology has ensured this. Technology and artifical anything centralizes all systems. This is fundamentally why centralized healthcare is simply gang warfare at scale for your healthspan and longevity. It has become so bad that now health influencers are replacing centralized MDs with these concepts and are now employing technology to allow them to quantify themselves against metrics they BELIEVE promote health based upon these centralized principles. They are codified in algorithms. They are trying to sell this idea to the public directly, without the Oracle problem of the centralized physician to block them. They are asking the public to reject the central dogmas in mediicne that have been developed in industrialized healthcare for the last 200 years. Ironically, the publis has no idea these people are making the same mistake as the physicians have. They are relying on a technocracy that used light incorrectly in their algorthms to spit out programs that make your health worse. This is why for 50 years every time humans have increased technology spending NHANES data shows our health gets worse. This is why we spend 4.3 trillon dollars a year on healthcare and we are the sickest nation on Earth. Our return on equity for our decisions making is horrendous because we have uncoupled the wsidom of nature from our ability to make dopamine and melatonin from sunlight we need to think well. Today people, who are sick and dying, realize that they can no longer afford to abdicate the decision process to someone else who embraces centralization. They no longer want to feel helpless. They want some degree of control back. This begins with embracing decentralized medicine. We advocate for optimizing life by advocating for effienct use of light, water, and magnetism to create dopamine and melatonin to increase the public’s ability to think and reason well to make better choices that expand healthspan and longevity and decouple from industrial healthcare process.
Central industrial medicine has left these concerns unaddressed for too long. The gap between recommendations and results is widening every day. Patients sense it and doctors feel is. This is why they are burning out and killing themselves at record rates now. We ignore it in hospitals and the centralized offices that now employ MDs and give them evidence based guidelines that fill bank accoutns and not our patients health accounts. Some of us recognize it and try things to close those gaps to remain competitive and up to date. Big companies and government will say they try to bridge these gaps with tangible action plans. Most of them are highly destructive to healthspan because of the incentives they built within the system. They are the carbon Oracle we aim to reduce in decentralized medicine. Most of the changes centralized entities have added to the system in the last 50 years have lead to incremental change that lead to spectacular declines in the public;s health under their watch. Centralizing forces in government allowed corporations to take over and this lead to regulatory capture. The people profiting from sick care now regulate the industrieds they control. More over, private equity, namely hospital systems, have bought doctors practices and turned them into employees who now have to act as their employer mandates. This make them the Oracle problem that stood between patient and doctors. We saw how spectacularly bad this was during the COVID epoch when we in a supposed pandemic and hospitals were firing the people trained to help people. They muzzled those of us who told people the right things to do because they were not good for the hospital’s profit and loss statement. Now we have data that the industrial military complex used cheap money and crooked scientists at the FDA and NAIAD to fund gain of function research that was used against the public. This system is off the rails and needs to be dismantaled. I have a plan to do just this.
The reason that success is often limited in centralized system is because they underestimate the personal behavioral aspects that often result in ultimate failure of their plans. My belief is that if we construct a decentralized paradigm of change that incorporates the tangible (costs and services) aspects of change and the best personal needs of patients needs then we will have the ability to effect change in both spheres simultaneously allowing for massive benefits that both sides of the equation. I have shared many of these ideas with politicians today.
My vision for healthcare is to focus on health promotion to allow people to adopt thoughts that immediately change their health with decentralized principles found in Nature. Organizational changes in the food agricultural complex will need to adapt with the current vertically and horizontally integrated healthcare delivery system. We need to adapt our service industry in healthcare much like the American government was able to morph our industrial complex in the 1940s to fight a World War but we must decoouple the industrial military complex from controling science or funding what should be studied in science. In WW2, that centralized campaign was successful because it allowed for change in both the tangible and intangible areas. The public and business sectors adapted for the betterment of citizens.
Today, the opposite is true. The military’s interventions into health has been a disaster for the public health. The best example of this is the botch distribution of the COVID jab by the DoD.
I believe my QUILT allows for this decentralized change in healthcare. My question is will you join me in that thought experiment? Will you become the patient or politician to help me change our current environment? We are human after all and we are perfectly capable of altering our environment are we not? My patients in my practice have been chasing change with me in this way for over 20 years. They understand that quantifying their healthcare using decentralized medicine is an experience is actually giving them the control back they seek.
TURING’s THEORY ON MORPHOGENESIS HAS NOW BEEN VALIDATED, BUT TODAY I AM UPDATING IT.
The chemical basis of morphogenesis is not the first step in life. Life begins with light. Light is transformed into electricity and all chemistry is preceded by a DC current generation in the cells of an organism. This idea has ancient origins of 3.8 billions years, as the slide below shows.
WHY DID I COME UP WITH THE IDEA TO USE YOUR CIRCLE OF SIX TO IMPROVE HEALTHSPAN?
It is suggested that a system of chemical substances, called morphogens, reacting together and diffusing through a tissue, is adequate to account for the main phenomena of morphogenesis. Such a system, although it may originally be quite homogeneous at its onset at the zygote level, may later develop a pattern or structure due to an instability of the homogeneous equilibrium, which is triggered off by random disturbances. In life’s cases, endogenous light generated by cells created an instability within your body plan soon after fertilization and this set up the dissipative state of humans. It sets the tone of what type of silly talking monkey will emerge from the placenta in 9 months.
Morphogenesis is a transgenerational epigenetic program born from light’s interaction with matter found in the parents germ lines. The human zygote is a reaction-diffusion system built around quantum computations. We should consider them each in some detail in the case of an isolated ring of cells, a mathematically convenient, though biologically unusual system because of the queerness that light contains. In quantum biology light investigates the matter contained within the germ line to give feedback on the current state of the environment. This implies that the germ cell line is chiefly concerned with the onset of instability to create the dissipative state life needs to evolve. It has been found since August 14, 1952 that there are six essentially different forms which this may take. Those six forms are the original building blocks buried at the core of the retino-hypothalamic tract that sculpts your body and your life. Turing’s idea on morphogenesis is where the circle of six idea came from in my own mind in 2005. All of these are linked to electric polarity changes that occur during morphogenesis.
Morphogenesis in biology, when you understand it is like watching a blind man try to solve a Rubik’s cube problem.
When you see embryology from two germ cells and then grow into an adult with a myriad of different cells you begin to see why timing became the key innovation for Mother Nature early on. She had to develop time crystal proteins to pulse via light waves to shorten the duration to innovate matter in living cells and then direct them how to migrate to the place of eventual function.
Truly a modern engineering problem if you think about the complexity mathematically.
The mathematical perspective for morphogenesis was given to us by Turing but the necessity for a mechanism of feedback interaction between universal structures given by Fred Hoyle in 1983 really makes sense.
Hoyle calculated the probabilities of a blind person ordering the scrambled faces of a Rubik cube. The calculations demonstrated that, due to the fact that the blind person does not know if he or she is getting closer or further to the objective on each move, the probabilities of matching the six colors on each face of the cube are on the order of 1:1 to 1: 5×10^18.
Thus, if that person was to labor at a rate of one move per second, it would take 5×10^18 seconds to complete all possibilities.
That is to say that it will take up to 158 billion years for that person to reach the goal.
Clearly, that time period not only grossly exceeds the life expectancy of the Rubik cube player, but it exceeds the lifetime of the Earth or for that matter the existence of our Universe since its estimated inception some 13.7 billion years ago. You can see Mother Nature had to solve this time problem really quickly.
So how did she do it?
Hoyle suggested some basic decision prompts.
If the blind person is given a simple piece of information, something like a “yes” or “no” prompt every time a move is made, which is every second, then the time needed to complete the Rubik cube equation is drastically reduced to two minutes.
I have a sense Mother Nature used the Aromatic amino acids in our cells to make the yes and no signal using ELF-UV pulses. The IR-A signals were used to keep the semiconductive powerplants functioning properly in the central retinal pathways to make these decisions.
This hints that our eyes hold a lot more power than we think for quantum computation.
The Retino-hypothalamic tract of the central retinal pathways is likely the root cause of 90-95% of modern diagnosed disease. This is not hyperbole, and can be actually proven by careful observation, just like the Rubik cube example above. From a neuroanatomy perspective if one lays out the entire structure and what this tract communicates with, it shows that it that allows light to touch directly or indirectly just about every tissue represented in the human brain. Moreover, the central retinal pathway is loaded woth more DHA and aromatic amino acids than any system in the human brain. This allows it to act as a semiconductive circuit that makes rapid decisions for many parts of the brain.
What does the semiconductive circuit do in the eyes?
It contains the SCN and the eye clock mechanism of the entire circadian system.
It contains 3 key semiconductors are DHA, melanin, and water.
it contains 2 semiconductors that regenerate the whole system: dopamine/melatonin
It’s also houses the root cause of your current mood and level of sleepiness.
It’s contains data on why you get hungry, your appetite/satieity and your energy status.
It controls all your pituitary outputs for hormones.
It controls all water balance in your body.
It controls endogenous melanin production in the brain
It links these two major semiconductive systems, water and melanin which generate energy and information processing in you
It is a gateway to your emotions in the frontal lobes and limbic system
It connects your new brain (frontal lobes) to the oldest parts of the brain (olfaction)
It’s also why houses the decision making apparatus that informs you, who you should listen too.
The light mediated rhodopsin system is activated by non visual red and UV light. Visible light is captured by the retina and delivered to a camera system and a vegetative system. The bookends of the visible spectrum feeds into the Rhodopsin mediated non visual, light system. A network of fiber optic cables branch from a special collagen at the start of the Rhodopsin tracts. I forget the protein name. It’s collagen that passes light the fastest The camera system allows you to see and run the clock, but the vegetative system informs the autonomic and non autonomic systems of the brain as well. Light makes this tract tick like a pure crystal with the cosmos. It defines what a quantum biology system should be. It is optical lattice clock and quantum computer working in parallel that alllows you to live life, all while being powered by a DC electric current. It houses the tissues in you, where light is FIRST polarized and the DC current modified for specific physiologic purposes of form and function. It is where the DC electric current becomes polarized for your stem cell depots to become a regenerative force in the body for every cell. This is why nothing should be worn in front of your eyes to ruin this optical signaling. This machinery allows your cells to dedifferentiate into more primitive cells to replace tissues damaged by the act of living on Earth. It is astounding what it is capable of.
The retino-hypothalmic tract is bidirectional, but during wakefulness it allows the input of photons to pass current in one direction to keep you conscious by quieting the default state of the brain in the brain stem. In DC currents the flows of electrons is from positive to negative, and this means electrons are actually moving opposite its normal direction. In this circuit, electrons flow from a scarcity region to where they become more abundant. This makes the signal unusual. Recall that electric field also form around any charged particle. The electromagnetic field environment around a DC current is very stable. The same is not true around an AC current. An AC current electromagnetic field collapses and reappears with its poles reversed every time the current changes direction. This reversal happens 60 times a second in the USA power grid. Magnetic fields around AC currents are very problematic in AC current environments. All current induces magnetic field. The AC magnetic field is variable. When this magnetic field moves in relation to the conductor, it induces a current. Any varying magnetic field, like that around tech gear or household appliances, generates stray currents in nearby conductors. Electric and magnetic field are represented by lines of forces and it is these forces that link to morphopgensis changes and physiologic abilities. Marsh and Beams experiments in planarians really elaborated these ideas in 1952. Their experiment showed that once could control the body plan of an animal via polarity changes. They should that they could control morphology by passing a current through it. Eventually this paper lead to work that showed us neuron tracts do orient themselves along a current’s flow. This told us Bernstein’s action potential hypothesis was flawed.
Consider Leduc in 1902. He used high voltage AC current to induce sleep in humans. Back then no understood why this happened but if you understand what I just said in the paragraph above you can see why it happened. Its changes the current, the charges, and the polariity of the field in the brain. This idea marries to all the experimental proof that Becker generates in his salamander regeneration experiments.
These tracts come out and take a round about route to final termination at the hypothalamus. All major brain nuclei have termination tracts from the retinohypthalmic axis. This axis contains the leptin melanocortin pathway and it innervates the entire neural territories that are adjacent to ventricular system that are loaded with CSF. CSF is a special semiconductor mostly made out of DDW and minerals to act as dopants. Even our motor function is impaired by alterations of this axis. Your motor behavior is also solar light mediated. This is why the light where you move and work out in is a critical part to get right.
It turns out you cannot do much unless you have proper variation of photonic frequencies from the visible spectrum powering the RHT. It kinda looks like the tract traces out a bow tie in our brains in 3D space and eloquently hits all 6 brain nuclei and terminates as an inversion at the hypothalamus. It’s morphologic design is a marvel of Nature. It’s morphology links to the type of electric and magnetic field incursions that occur here.
It has its own resonate frequency that travels along optical chasms and reaches every part of the body. This tract uses melanin and water semiconduction as a quantum information lattice. It uses the ration of H+ to D and the number of electrons to run its entire program of decision making. I covered those basics in the Kruse for Dummies lecture.
How might hve Mother Nature have solved this thermodynamic puzzle of life?
I looked back to the museum for clues.
Fred Hoyle, origninally was a mathematician of Cambridge University and presented his theory known as ‘supernova hypothesis‘ in the year 1946. His hypothesis was based on the principles of ‘nuclear physics’ and was described in his essay entitled ‘Nature of the Universe’. From this theory came the ideas of the Big bang and how stars made elements from exploding stars. Light was behind this trick.
There was another trick I learned about light as a boy in New York City.
The PHOTOELECTRIC EFFECT.
I read a lot papers on UV light as a resident in neurosurgery and I knew a lot about UV light and aromatic amino acids in the brain. An important photochemical mechanism that occurs using the photoelectric effect in proteins involves reduction of disulfide bridges (SS) upon UV excitation of Tryptophan and Tyrosine side chains (Kerwin & Rammele, 2007, Neves-Petersen et al., 2002 & 2009a).
UV-excitation of tryptophan or tyrosine can result in their photoionization and to the generation of solvated electrons. The generated solvated electrons can subsequently undergo fast geminate recombination with their parent molecule, or they can be captured by electrophilic species like molecular oxygen (02).
Either option here could have been the yes or no answer mentioned by Hoyle’s work on how stars create matter from light.
Molecular oxygen decreases the chance of pseudohypoxia and this make formation of an exclusion zone (EZ) in water in skin cells. This leads to coherent domains within water, (at a lower pH than normal) that allows photo molecular interactions to occur between cysteine and cystines. You can go read my Energy & Epigenetics #12 blog to see where I dropped that breadcrumb long ago.
In different tissues, in the case where the electron is captured by the cysteine, the result can also be the breakage of the disulfide bridge in proteins like glutathione (Hoffman & Hayon, 1972). I’ve mentioned this in the Energy and Epigenetics 12 blog.
Here you begin to see how full spectrum UVA light and the aromatic AA begin to work to create energies and information that can modify sulfhydryl groups in proteins and lipids in different tissues to control non-linear optical gating of signals from sunlight’s frequencies to control morphogenesis. Remember, sunlight sulfates many proteins to make them water soluble so that electrons in water become useful to cells and tissues. Melanin unlocks those electrons moves freely within the retino-hypothalmic system. Melanin absorbs all light frequencies to liberate massive amounts of electrons to be used in semiconductive proteins in the systems of biochemistry. I like to think about biochemistry as a block chain and I think about the DC electric current as the PoW mechanism that powers it. The hash rate of the system is controlled by melanin’s efficiency at liberating electrons for the biochemical blockchain inside of our quantum computer. This is how we have to crack the ENIGMA CODE of biology.
Activated excited electrons from the incident UV light is where the process begins. We also know that biophoton creation in numerous locations in the cell always needs oxygen and ROS/RNS to be present for light to be made from Roeland van Wijk’s book that has an amazing bibliography of the science of endogenous biophoton creation by cells.
This is purely a photoelectric process and it used for timing processes critical in the morphogenesis of our organs. This is how life sculpts life and how it built us. Turing’s paper in 1952 gave me the idea of how the process could be described mathematically but no idea of how biology completes this task. I am attempting to do that right here and right now with my words which are sculpting these ideas.
The UV light collisions are also created thiol free radicals in this process with the amino acids with disulfide bridges. Those radicals can build or subtract the overall quantum spin state of the differentiating cell or dial in a key Orbital Angular Momentum number contained in each light photon we generate and a cell needs to sculpt a certain tissue from a homogenous stem cell in a zygote. All of this information could easily be stored magnetically in DNA. This is the basis of the ideas that are contained in the Kruse for Dummies lecture. How the Rosetta Stone works in biology was not how Darwin wrote the rules.
Never forget that light creates the matter in the universe when the light is manipulated in some way physically by matter. Timing controls how light energy controls the matter in cells. You are made of tissues that are filled with mitochondria, which generate light inside of you that control this process. They harvest parts of atoms from foods you consume, tear them apart and decipher an ancient code in them like a blockchain does. This is essentially what biochemistry is for a quantized system. Biochemistry is a blockchain factory used to make things a tissue needs using spin state and OAM alphabets in the mitochondrial code of life.
Throughout 4.5 billion years of molecular evolution, proteins have evolved in order to maintain the spatial proximity between aromatic residues (Trp, Tyr and Phe) and disulfide bridges (SS) (Petersen et al, 1999).
We might call this mechanism of how light interacts with certain aromatic amino acids and sulfur-containing amino acids how the fractal geometry of life begins.
There is a very special spatial geometric relationship that exists because the process is quantized to light frequencies that our star releases to us on Earth wirelessly. This has not been well appreciated by modern centralized healthcare. This is also why the sun’s light reduces all-cause mortality and why it can never be replaced in Big pharma healthcare. I also believe this is why solar redox is critical to having a child with great morphogenesis.
To suggest this is lunacy when you understand biophysics well.
The interaction of the most powerful part of the solar spectrum of light (UVA/B/C) measures the collisions in the aromatic amino acids and in the disulfide bridges. The aromatic amino acids location becomes the first step in determining where the position and geometry of residues to act as nanosized antennas in the protein world that can capture UV light (from ~250-298nm). This could be other yes or no binary codes cells use to build tissues in the Rubik’s cube example above. You can see why the idea of a time crystal is paramount now when you are building a life form. Think about my Vermont 2018 video now in this light!!!
The first two protein bends are always determined by nuclear DNA coding and this would save us a lot of time in morphogenesis. The last two bends are tied to the redox state which is linked to this quantum photoelectric process I am describing here now. If the timing of the yes and no’s are off the protein folding is off, and you’ll make glitches in the folding of proteins and organelles in cells and tissues.
Once excited by the incident ELF-UV light these amino acids can enter photochemical pathways likely to have harmful or beneficial effects on protein structures (yes and no again) by affecting specific bonds like disulfide bonds in cysteine/cystine. In the circadian mechanism, their effect on the PER protein that is made every AM by sunlight exposure on our body is critical in the right amount of phosphorylation in our cells to tell time.
These two amino acids are the rarest amino acids in our proteins, and as such can acts as the ideal photo-optical switch or gate for signaling because of they a relatively rare in humans.
It turns out cysteine/cystine disulfide bridges in proteins are known to be excellent quenchers of the excited state of aromatic residues by UV light in the literature. This means these disulfide residues naturally decrease the power present in UV light created in the nearby excited aromatic amino acids of the cells in a developing life form.
In this way, they contribute to protein stability and activity in tissues, thereby, stabilizing ubiquitin rates and helping drive morphogenesis via the blueprint in the OAM of DNA. I assume each species has its own molecular resonance OAM they operate with.
UV light excitation of the aromatic residues is known to trigger electron ejection from their side chains (Bent & Hayon, 1975a; Bent & Hayon, 1975b; Bent & Hayon, 1975c; Creed, 1984a; Creed, 1984b; Kerwin & Rammele, 2007, Neves-Petersen et al., 2009a).
These electrons can be captured by disulfide bridges in simple proteins with tons of disulfide bridges like glutathione, leading to the formation of a transient disulfide electron adduct radicals, which will dissociate photoelectrically, leading to the formation of free thiol groups in the protein. what you did not know until my Patreon blogs in recent years is that DDW water created by the mitochondrial matrix can increase endogenous glutathione for matter creation and organ building.
This photochemical change then leads to non-optical signaling at deeper levels in the embryo as development continues using the butterfly effect. When this process is inhibited in any way, a transgenerational disease manifests in a fetus or child. This is what autism, cancer, or a congential anomaly is at its core level. It is a blockchain that causes a hard fork in the offspring.
The irony in all these details is that UV frequencies foster the creation of matter naturally, and do not cause cancer, by these mechanisms.
More irony for centralized healthcare myopic paradigm that the sun and UV light is bad: This mechanism is now being used big pharma to develop drugs using nanotechnology and the ability of cells to make ELF-UV light in a process called LUMI.
THE KEY INSIGHT LEARNED ABOUT LIGHT HAPPENED WHEN I WAS 5th GRADE ON A SCHOOL TRIP.
I found out we had an incredible ratio of light photons compared to atoms in the Universe.
How would all this lead to logic theory based on computation? This juxtaposition might be fun and a bit shocking. You have to understand some basics about information theory to make sense of Turing was doing in his brilliant mind.
Biologic researchers are paying much more attention LATELY to mechanisms that “actively erase” and hide memory engrams in the brain.
WHY would they do this? If you cannot erase memory you cannot build a life that is based upon thousands of quantum computers working in unison to skate around the second law of thermodynamics to become the ultimate Maxwell demon.
What do I mean?
The erasure of information is what drives entropy increases over time. This is a consequence of having 1 billion light photons to one atom ratio in the universe. All molecular clocks that control circadian biology are, are flow meters for entropy. This told me circadian biology was the key to the biological ENIGMA CODE of all modern diseases.
Because of information theory the brain has to be designed to forget information on purpose to satisfy the dictims in the second law that Maxwell hinted at in his letter to a friend written about “demons”. We later called them Maxwell demons.
The first law of thermodynamics says energy is fixed in the cosmos because it cannot be created or destroyed. If energy is fixed in the universe, and the speed of light is fixed in a vacuum, what does it mean when we know there are one billion photons present in the cosmos for every atom in the known universe?
It means light is the “Jacquard card” or the Claude Shannon punch card of life inside of cells……..
Jacquard built the first computer (pre-Shannon) that used punch cards to make silk patterns using a binary pattern of open circles and closed circles using the loon. Shannon did the exact same thing for Bell Labs using the binary code.
So how did Mother nature build her first computer called a cell? She used light.
Cells are all quantum computers filled with biological semiconductors. They do not look like the objects found in your cell phone.
Well because there are one billion photons for every atom she started with light. She realized 4.6 billion years ago that light contains both energy and information at a ratio of a billion to one. Of the two she was the first to realize that information in light could be used to organize matter.
It took until Maxwell’s paper on Maxwell demons in the 1860s, for anyone else to realize this.
Light is the qu”bit” cells use to transfer information. Information is buried in Orbital Angular Momentum of photons in the Universe. They are ubiquitous. The energy in light is buried in frequency and a whole lot more of the characteristics linked to the photon. Since there are one billion photons per atom energy and information has to flow from light to atoms. It cannot flow the other way, just like a waterfall cannot flow in reverse.
A billion to one ratio is like asking a 20,000 foot waterfall to reverse its flow without adding a bit of energy to it. IMPOSSIBLE.
WHY ARE CENTRALIZED MEETINGS OF SCIENTISTS ABOUT AS USEFUL AS THE PICTURE BELOW?
DID THIS IDEA HAVE OTHER MASSIVE IMPLICATIONS?
Yep. It explains why sleep exists and why it is restorative to healthspan.
Information can create order from chaos only when we store information about light in our memory (water). What is the physical basis of this? With elapsed time, just as RAM or a hard drive becomes full, light information will fill the capacity of water’s hydrogen bonding network and fill up all the possible memory, and memory will then become a drag on function and life will decline. How did life change this phenomenon? It innovated sleep. Sleep is when we prune our memory in light and in water. Our liquid crystalline arrays have to be deleted every night. Information can only be useful on an ongoing basis if it is useful and used over and over and deemed critical, or until it is deemed useless and the memory mechanism in cells deletes information contained in light or water from our stores. When this happens it balances the second law of thermodynamics. It appears life breaks the law but she never does because she return useless information back to the environment she no longer needs to build order and the living state.
This means there really is no free lunch in living things. How did life do this? Evolution built sleep to be mandatory so it could delete data at night when light was absent. If it is not absent you cannot delete the data and your clocks timing mechanism is DEFECTIVE in this tissue. This leads to disease. This is where memory in water is being deleted diurnally so that we can continue to use the information to order our cells while we live.
SUMMARY
The erasure of information is what drives entropy increases over time. This is a consequence of having 1 billion light photons to one atom ratio in the universe. Light is turned into current easily because of this relationship by matter in cells.
We see this in the work of John von Neumann. Like Turing, he was also a mathematician.
In analog computers, changes in information are expressed in analogous changes in the magnetude or polarity of currents. A computer is used to store the varying temperatures of a furnace, the rise and fall in heat can be mimicked by a rise and fall in voltage.
Analog systems are slow and can handle simply information but they can express subtle variations very well. Digital coding, on the other hand, can transmit enormous amounts of data at high-speed, but only if the information can be reduced to a yes or no, on or off bits of information like 1 and 0.
The human nervous has been found to be a hybrid computer. We now know analog coding controls the overall activity of large groups of neurons by such actions by increasing or decreasing their sensitivity of incoming messages.
The digital system would transfer sensory and motor information, but the processing of that information-memory and recall, thinking, and consciousness would be accomplished by the synergism of both methods. Awake brain surgery proved this is exactly what goes on in the human brain when we are simultaneous checking electrical activity of the cortex in an awake and anesthetized patient.
Water carries light information in the coherent domains of water’s hydrogen bonding network of pentagrams or hexagons. This is why water has the ability to carry a memory of lights OAM fingerprint. It is codified in the coherent domains of hydrogen bonds and its increasing dielectric constant. And we are designed to sleep every night to erase part of the information stored in water that cells find superfluous on a charge basis. (Luc Montagnier)
What are the laws of Nature telling us that we are missing in centralized medicine? If you listened to my recent podcast with Dr. Cowan you heard about the silicon dioxide crystals in Cite 5. How crystals transfer information to water has been a mystery to centralized science. Below you’ll see the picture of Montagnier 2009 experiment. But in Cite 5 you’ll see the science that underpins what Montagnier did from Marcel Vogel, and IBM scientist who worked peripherally in the MKUltra program.
If you can measure it, it’s not sacred to healthspan or life. What we delete alters the balance of what is being measured. Therefore it is superfluous. This is counter to how we think about the world. That is the queerness embedded in quantum mechanics.
Water carries light information in the coherent domains of water’s hydrogen bonding network of pentagrams or hexagons. This is why water has the ability to carry a memory of lights OAM fingerprint. It is codified in the EZ hydrogen bonds and its increasing dielectric constant. And we are designed to sleep every night to erase part of the information stored in water that cells find superfluous on a charge basis. (Luc Montagnier)
How do we delete information? Have you ever heard of the Landauer’s limit, because this is how cells do it?
Landauer’s principle establishes an irreversible conversion from information to physics. This Landauer principle desribes how thermodynamic entropy is converted into information entropy at the expense of negentropy also knows as “negative entropy” in a cell. You can read about it in CITE 3 below.
This is how the journey back home to the truth began.
Culture and creativity integrate wonder and wisdom. It is at the Intersection of Art and Bio-physics
Today, what we love we have become addicted to, and it has fueled our myopia to the truth.
Today’s thought experiment: LIGHT WATER AND MAGNETISM
ARE HUMANS BEHAVING LIKE A CANCER ON EARTH because of the light they use to communicate and live???
Is the evidence found in civilization all around us?
Cancer is when a cell from the human body errs in such a way that it:
1. Starts to replicate without control
2. Invades other bodily systems without care (metastasis)
3. Spawns clones within the same tumor, and these clones compete with each other to maintain tumor dominance (intra-tumor heterogeneity)
4. Clones that make it to another bodily system may be different from the primary tumor (inter-tumor heterogeneity)
5. Siphon off all energy/info sources to serve the needs of the tumor and not the body
6. Grow and grow and grow without respect for the borders within the human body (local invasion)
7. Grow new blood vessels to feed the tumor instead of the healthy cells (angiogenesis)
Humans brains shrunk 65,000 years ago. This was linked to melanin destruction as they left Africa and headed to the 51st latitude, where new speciation began inside caves powered by campfires. Art and creativity were the first evidence of what melanin destruction can do. Today, we see this as a positive. Initially, it might have been. But what are the collateral effects of this effect of light on our biology? Is it not unreasonable to say humans have become a cancer on Earth the more they have innovated uses of the electromagnetic spectrum?
Might they be manufacturing the 6th extinction and keeping you in the dark by removing all the fire alarms in society to achieve the goal?
Neanderthal brains shrunk with time as their light stability changed, morphing from equatorial light to light in the tundra.
Art and creativity were its atavist effect. This implies that art has profound lessons to teach us about health.
As light stability was lost and metabolically expensive, larger brains became costly.
Magnetochemistry and POMC sculpted the skulls of our ancestors. Genes had very little to do with these changes.
HISTORY OF MAN:
The human brain has nearly quadrupled in size in the six million years since our species last shared a common ancestor with chimpanzees.
However, studies show this trend toward larger brains has reversed in Homo sapiens. Our species’ average brain sizes have shrunk over the last 100,000 years.
For example, in a recent 2023 study, he tracked the braincase volumes of ancient hominins on a spaceship through time. He started with the oldest known species and ended with modern humans.
They found that rapid brain expansion occurred independently in different species of hominins and at other times across Asia, Europe, and Africa. Species whose brains grew over time include Australopithecus afarensis, Homo erectus,Homo heidelbergensis, and Homo neanderthalensis. It stopped with them.
Why?
The light they lived under changed. Equatorial light has light stability. High latitude light does not have light stability.
The trend for brain enlargement over time was turned on its head with the arrival of modern humans. The skulls of men and women today are, on average, 12.7% smaller than that of Homo sapiens who lived during the last ice age, as the picture above shows.
Note for those who follow culture: When our ancestors had larger brains, there were no culture wars because there was no culture present in the history of homo species up until this time.
So, how can we explain this striking reduction? A study of paleoanthropology suggests that the shrinkage in brain size began around 100,000 years ago when the last ice age began. This corresponds to a period in which humans switched cognitive abilities because this was the time when humans began producing symbolic artifacts and engravings with meaningful geometric images on them. These were the precursors of artwork that was soon to appear on the walls of caves.
As they left the cradle of humanity, temperatures changed, light stability was lost, and more glucose was made from high-latitude light. That higher glucose level created a relative pseudohypoxia that created the need to reduce the size of the brain thermodynamically. The human brain shrank due to the abscopal effects of blue light from a latitude change and the need to use fire to keep warm. This fire was used inside a cave, and for the first time, humans began to use animal skins to cover their skins. The skin is a solar panel for the brain, and with it covered, brain expansion was halted dead in its tracks.
Smaller brains have allowed humans to cool down quickly. It’s well known that humans in hot climates have darker skin and more melanin and have evolved leaner and taller bodies to maximize heat loss. Warmer temperatures link directly to equatorial light stability and melanin biology.
DID YOU KNOW CREATIVITY AND LANGUAGE BOTH SHOWED UP WHEN SUNLIGHT WEAKENED IN OUR FAMILY? IS IT HAPPENING AGAIN IN OUR MODERN WORLD? IS THE RESULT REFLECTED IN ART AND CULTURE?
As man left equatorial Africa and entered the Middle East, the invention of written languages like Sumerian began
Nobody doubts that Albert Einstein had a brilliant mind. But the Nobel prizewinner, famous for his special and general relativity theories, wasn’t blessed with a big brain. Based on the history of hominin speciation, it was smaller than average.
This seems surprising to centralized science because they believe bigger is better. Quantum thermodynamics tells us that smaller is more energy efficient. This means the impetus to shrinkage is likely tied to energy and information loss from the environment. This story should have brought centralized science to the story of light, but it has not. Look at the picture slide show in this blog. The answer is here.
Big brains are a defining feature of human anatomy that we are proud of. Other species might be speedy or powerful, but we thrive using the ingenuity that comes with a large brain. Or so we tell ourselves. Einstein’s brain hints that the story isn’t simple – recent fossil discoveries confirm this. Over the past two decades, we have learned that small-brained hominin species survived on Earth long after big-brained ones appeared. Moreover, evidence shows that they were behaviorally sophisticated as the brain shrunk. Some, for instance, made complex stone tools that could probably only have been fashioned by individuals with language.
These discoveries turn the question of human brain evolution on its head. “Why would selection favor big brains when small-brained humans can survive on the landscape? Neural tissue consumes lots of energy, so big brains must have benefitted the few species that evolved them. But what?
The answer is that it allowed us to concentrate melanin inside our skulls because the light on the exterior created so much POMC translation at our surfaces. The more melanin created on our skin, the more it allowed it to move to our interiors, where it became housed endogenously as our most important wide-band gapped semiconductor that sculpted our body plan from our cousins. The result: Humans have more cognitive abilities than any other primate in history.
So what happened as the brain shrunk? Melanin was degraded and lost, and 125 grams of neural tissues were subtracted, but the amount of CSF in the human brain increased relative to its size.
WATER BECAME MORE IMPORTANT TO THE BRAIN FOR FUNCTIONING.
WHAT IS THE ART CORRELATE TO THIS IN HUMANITY?
At the end of the line, the aqueduct’s contribution to modern human flourishing is marked with a nod to the other great civilizational force that built Rome:
Beauty.
Meet the Trevi fountain. It could have never been built by a Neanderthal brain.
Just as the functioning of the human brain improved when neurons shrunk and water increased, so did the Roman Empire.
At their peak, these systems fed Rome with a million cubic meters of water per day, which was used for baths, fountains, and sewers.
Clean water was the civilizational lifeblood that allowed Rome and its population to rise above all others.
How are stars and water linked?
WHAT ELSE CHANGES THE REFRACTIVE INDEX OF A SUBSTANCE EMITTED BY STARS WHEN THEY ARE CREATED?
WATER.
So, was the shrinking human brain filled with more water and somehow fossilized into human culture?
I think so.
The Trevi Fountain
What’s behind is often forgotten: a 17th-century palace called the Palazzo Poli.
Salvi turned its entire facade into his great stone symphony carved into rock. But people often miss something far more interesting: what’s underneath.
It’s the endpoint of the last of Rome’s 11 ancient aqueducts, the Aqua Virgo, which is still in operation.
The fountain was only completed in 1762, but it was built on a site of ancient significance. This is the same idea we see in the cenote system in Mexico with the Maya.
Following the water beneath the fountain, you’ll find the Vicus Caprarius — the City of Water.
It’s a first-century housing complex, and the water flowing through it and the Trevi Fountain’s water still comes from the ancient aqueduct.
If you follow it further (red line), you’ll end up 20km outside of Rome at the springs that feed it.
Just as water was the key to power generation in the shrinking human brain, Rome’s aqueducts were the key to its dominance: remarkably simple in concept and brilliant in execution.
To bring fresh water right into Rome, all you needed was a slight, continuous gradient from source to endpoint, and gravity handled the rest.
That meant digging tunnels to cut straight through hillsides and erecting bridges to cross valleys…
Sometimes, it meant building 160 feet into the air. From one end of the Pont du Gard to the other (900 feet), the level descends less than 1 inch. The Roman Empire was built on impeccable precision.
At their peak, these systems fed Rome a million cubic meters of water per day: baths, fountains, sewers… Clean water was the civilizational lifeblood that allowed Rome and its population to rise above all others.
At the end of the line, the aqueduct’s contribution to human flourishing is marked with a nod to the other great civilizational force that built Rome:
Beauty.
MAGNETISM
When melanin degrades, there is less of it to deal with the magnetic chemicals made by mitochondrial metabolism. Those chemicals are ROS and RNS. These chemicals control metabolic flux in a cell. Stochastically, they all link back to the light we live under as well. This is why all metabolic pathways have different levels of ATP associated with their use.
Changes in ATP always link to ROS and RNS production, which is why we did not need genes to create man. ROS and RNS chemicals all have one unpaired electron, which makes them the source of magnetic flux in cells. Magnetic flux controls timing sequence in cells and timing is the key to morphologic changes in evolutionary creation. The same is true in our brains concerning art creation.
We used changing light frequencies to become who we are. Our identity is reflected in our art, which begins in our hydrated carbon-based semiconductors. See the paper written three days ago. That is how cutting-edge the science of Nature is.
Diamonds are carbon-based semiconductors.
Did you know that diamonds don’t actually shine on their own?
They reflect light the semiconductor emits!
Might the hydrated carbon-based semiconductors be reflecting their magnetic effects in the artwork humans create? Art is built from that reflected light in our brains.
I think that is exactly why visionary artists use quantum entanglement with their environment to become the first members of a culture to see the world in a new way. This is reflected in their work. When it manifests in reality, culture flows to the rest of the silly-talking monkeys.
This fascinating property is one of the reasons why diamonds make excellent semiconductors.
Here’s why:
1. *Crystal Structure*: Diamonds have a unique tetrahedral crystal structure that allows for efficient electron movement, enhancing their performance as semiconductors. Water made by metabolism also has its own unique geometry.
2. *High Thermal Conductivity*: Diamonds can dissipate heat effectively, crucial for electronic devices to function efficiently and reduce overheating. The water around our semiconductors allows for the same thing.
3. *Wide Bandgap*: Diamonds have a wide bandgap, making them ideal for high-voltage and high-frequency applications. This enables more efficient electronic devices and systems. Diamonds and all hydrated carbon-based semiconductors work well with UV light. I have established in the Quantum Engineering series that we are filled to the gills with wide-band gapped semiconductors that use UV light.
4. *Chemical Stability*: Diamonds are chemically inert, which means they can function reliably in harsh environments. So, the next time you admire a diamond’s brilliance, remember that it’s not just about beauty but cutting-edge technology! Our semiconductors are inert as well under the power of visible light. Any other frequency in the electromagnetic spectrum induces changes to our semiconductors.
Our use of the light spectrum is why chronic diseases are upon us. We have the same number of genes as our nearest relatives. Few see what I see. After I have infected your mind with this idea, I want you to visit any museum and see the history of art.
“What would happen,“Marshall McLuhan wondered in his seminal 1964 treatise Understanding Media: The Extensions of Man, “if art were suddenly seen for what it is, namely, exact information of how to rearrange one’s psyche to anticipate the next blow from our own extended faculties?
A surgeon is both an artist and a scientist… Surgeons rely heavily on their intuitive visual-spatial right-hemispheric mode. At the same time, our training is obviously scientific. Left-brained logic, reason, and abstract thinking are the stepping stones leading to the vast scientific literature’s arcane tenets. The need in my profession to shuttle back and forth constantly between these two complementary functions of the human psyche This ability has served me well to see the unknown and hidden messages of art history and evolution.
These thoughts today are on the quantum biology of farming. I show you what planting seen inthe public’s fertile mind can accomplish. You’ll never look at art the same way again. Humanity creates beauty when it is entangled with Nature. It establishes simplified things when it is made to the wrong frequencies of light. Centralization results from this prescription.
Don’t fear burning down art critics’ beliefs about their world today. Today’s ashes are tomorrow’s soil. Moments, like my talk at Palestra Society on Friday, August 15th, 2024, are like seeds to place in that soil.
Any moment’s beginning ends in a moment….and the seed transforms into something new, something different, something unique. The moral of the story of the seed is that if you want something new, you must stop doing something old.
This view below of the sun says something significant about the modern environment of man in the 1960s (above pic)
We have made Nature artificial, and it is reflected in the painting.
Art and the physics of quantum biology are propelled by revolutionary insight — that transcendent clarity of vision that Rilke called a “conflagration of clear sight” — which reframes our understanding of the world.
Although the development of physics has always depended upon the incremental contributions of many original and dedicated workers, on a few occasions in history, one physicist has had an insight of such import that it led to a revision in his whole society’s concept of reality. . . .Think about the contributions of Newton, Faraday, and Einstein as examples.
Emile Zola’s definition of art: “Nature as seen through a temperament,” invokes physics, which is likewise involved with Nature. The Greek word physis means “Nature.”… Like any scientist, the physicist sets out to break “nature” down into its component parts to analyze the relationship of those parts. This process is principally one of reduction.
On the other hand, the artist often juxtaposes different features of reality and synthesizes them so that upon completion, the whole work is greater than the sum of its parts. There is considerable crossover in the technique used by both. The novelist Vladimir Nabokov wrote, “There is no science without fancy and no art without facts.”
In addition to illuminating, imitating, and interpreting reality … artists create a language of symbols for things for which there are yet to be words. Their works are reflections of the light they are forced to imbibe. And we consume it. It reminds us of us we cannot put our finger on, but we know there is a deep connection. This series of blogs makes concrete decentralized connections.
SUMMARY
Art lives not only as an aesthetic that can be pleasing to the eye but also as a distant early warning system occurring in our colony of mitochondria and the collective thinking of a society.
Visionary art alerts the other members that a conceptual shift is about to occur in the thought system used to perceive the world. For the last 120 years, I have not seen one DaVinci, Rembrandt, or Monet producing among us.
In my opinion, Art creation has been creatively dying since 1905.
Is this related to when Marconi harnessed the power of light in radio and wireless transmission?
I’ll leave you to ponder this.
My vision is based on art critic Robert Hughes’s assertion that “the truly significant work of art is the one that prepares the future” and adds:
Repeatedly throughout history, the artist introduces symbols and icons that, in retrospect, prove to have been avant-garde for the thought patterns of a scientific age yet to be born.
They are born; they are just random packets of quanta awaiting to be organized by the melanin in your mind.
Decentralized science carries the torch of a tradition that stretches back through a long line of rebellious thinkers who have resisted the usual dividing lines between physics and philosophy. It is in this space where art is explanatory to human evolutionary history. In experimental metaphysics, the tools of science can be used to test our philosophical worldviews, which in turn can be used to better understand science that remains fuzzy to centralized thinkers.
Metaphysics is the branch of philosophy that deals in the deep scaffolding of the world: the Nature of space, time, causation and ex,istence, the foundations of reality itself. It’s generally considered untestable, so centralized science ignores it. This is done at the peril of the public health. Since, metaphysical assumptions underlie all our efforts to conduct tests and interpret results. Those assumptions usually go unspoken.
Most of the time, that’s fine. Within centralized intuitions they foster beliefs of how the world works for their benefit, and it often conflicts with our everyday experience of reality. At speeds far slower than the speed of light or at scales far more significant than the quantum one, we can, for instance, assume that objects have definite features independent of our measurements, that we all share a universal space and time, that a fact for one of us is a fact for all. As long as our philosophy works, it lurks undetected in the background, leading us to believe that centralized science is something separable from metaphysics mistakenly.
But at the uncharted edges of experience, where the quantum biology of the brain lies — at high speeds and tiny scales — those intuitions cease to serve us, making it impossible for us to do science without confronting our philosophical assumptions head-on. Suddenly, we find ourselves in a place where science and philosophy can no longer be neatly distinguished. Art and creativity of society can and do provide answers to the hard questions of evolution.
THE DIVIDING LINE between science and philosophy has never been clear. Often, it’s drawn along testability. Any science that deserves its name is said to be vulnerable to tests that can falsify it, while philosophy aims for pristine truths that hover somewhere beyond the grubby reach of experiments. So long as that distinction is in play, physicists believe they can get on with the messy business of “real science” and leave the philosophers in their armchairs, stroking their chins.
As it turns out, though, the testability distinction doesn’t hold. Philosophers have long known that it’s impossible to prove a hypothesis. (No matter how many white swans you see, the next one could be black.) That’s why Karl Popper famously said that a statement is only scientific if it’s falsifiable — if we can’t prove it, we can at least try to disprove it.
In 1906, French physicist Pierre Duhem showed that falsifying a single hypothesis is impossible. This should teach us that the basis of the scientific method has some limitations. Rarely do we talk about this in today’s science, but it was the talk of the day in the 19th and 20th centuries.
He argued that every piece of science is bound up in a tangled mesh of assumptions. These assumptions are about everything from underlying physical laws to the workings of specific measurement devices. If the result of your experiment appears to disprove your hypothesis, you can always account for the data by tweaking one of your assumptions while leaving your hypothesis intact.
Take, for instance, the geometry of space-time. Immanuel Kant, the 18th-century philosopher, declared that the properties of space and time are not empirical questions. He thought not only that the geometry of space was necessarily Euclidean, meaning that a triangle’s interior angles add up to 180 degrees, but that this fact had to be “the basis of any future metaphysics.” According to Kant, it wasn’t empirically testable because it provided the very framework within which we understand how our tests work in the first place.
Yet, in 1919, when astronomers measured the path of distant starlight skirting the gravitational influence of the sun, they found that the geometry of space wasn’t Euclidean after all—it was warped by the effect of gravity from stars, as Albert Einstein had predicted.
Or did they? Henri Poincaré, the French polymath, offered up an intriguing thought experiment. Imagine that the universe is a giant disk that conforms to Euclidean geometry but whose physical laws include the following: The disk is hottest in the middle and coldest at the edge, with the temperature falling in proportion to the square of the distance from the center.
Moreover, this universe features a refractive index — a measurement of how light rays bend — inversely proportional to the temperature. In such a universe, rulers and yardsticks would never be straight (solid objects would expand and shrink with the temperature gradient), while the refractive index would make light rays appear to travel in curves rather than lines. As a result, any attempt to measure the geometry of the space — say, by adding up the angles of a triangle — would lead one to believe that the space was non-Euclidean.
Any test of geometry requires you to assume specific laws of physics, while any test of those laws of physics requires you to assume geometry. Sure, the disk world’s physical laws seem ad hoc, but so are Euclid’s axioms. “Poincaré, in my opinion, is right,” Einstein said in a 1921 lecture.
He added, “Only the sum of geometry and physical laws is subject to experimental verification.” As the American logician Willard V. O. Quine put it, “The unit of empirical significance” — the thing that’s testable — “is the whole of science.”
The most straightforward observation (that the sky is blue or the particle is there) should force us to question everything we know about the universe’s workings.
But actually, it’s worse than that from a decentralized perspective. The unit of empirical significance is a combination of science and philosophy. The thinker who saw this most clearly was the 20th-century Swiss mathematician Ferdinand Gonseth. For Gonseth, science and metaphysics are always in constant conversation, with metaphysics providing the foundations on which science operates, science providing evidence that forces metaphysics to revise those foundations, and the two together adapting and changing like a living, breathing organism. As he said in a symposium he attended in Einstein’s honor, “Science and philosophy form a single whole.”
Revolutionary art has served the function of preparing the future at all times.
The art of today has me worried about our species’ future. We are headed back to our times in caves.
The art revolution today is being heavily influenced by artificial Intelligence. This concerns me, and it should concern you. Anything crafted by AI adopts a centralizing force in its DNA. This centralizing force sits at odds with the decentralizing forces in Nature.
Revolutionary art and visionary physics attempt to speak about matters that do not yet have words. That is why people outside their fields could better understand their languages. Because they both speak of what is to come, we must learn to understand them.
My art is quantum biology.
Our cognitive de-evolution is real, and it has been reflected in our art since Picasso.
We don’t see things as they are… we see things as they are.
There is a reason we see a lot of minimalist “art,” abstract modern painting, container houses, and buildings that are blocks of concrete with holes for windows when, in reality, everything is artless.
The minimalist trend is an ideology/formatting for people to accept less than they deserve, things that are unbalanced, artless, and heartless. If we look at things from a few decades ago, everything was ornamental, with style and thoughtful design. This shows what you are referring to: when society gets ill, that gets reflected in everything it creates.
Ask yourself right now who the 20-21st century great artists are.
Can you name anyone?
I cannot.
It even shows up in buildings when Marconi and Tesla were ruining humanity.
Some will look at a plumeria flower (gardenia/magnolia) and try to tell us that it results from perfection in minimalism.
This uncovers what they do not know. It is false. For me, it is a reflection of the melanin sheets in them.
The Plumeria flower and the Trevi fountain represent opposite poles of the complexity argument.
Did you know flowering plants like the plumeria species occur due to a lack of CO2 energy? So, their analogy is poor. They are minimalistic flowers/plants because of their low-energy environment. They are great examples of my point.
Plants also are made of DC electric semiconductors like we are. They reflect the light in their environment, structure, and phylogeny. Most plant groups were relatively unscathed by the Permo-Triassic extinction event, although the structures of communities changed. This may have set the scene for the appearance of the flowering plants in the Triassic (~200 million years ago) and their later diversification in the Cretaceous and Paleogene. The latest major group of plants to evolve were the grasses, which became important in the mid-Paleogene from around 40 million years ago. The grasses, as well as many other groups, evolved new mechanisms of metabolism to survive the low CO2 environments linked to warm, dry conditions of the tropics over the last 10 million years. Beauty varies as environmental energies vary. The results are in the art we create, the buildings we make, the brands we’ve built, and the flowers that do best as continue to spread artificial light over the surface of Earth like a viral pathogen spreading cancer.
At a certain point, you would think more people would realize that these minimalist cities are filled with antennas and nnEMF: They have accepted to be a total debt slave to the fiat system, and they do not seem to understand why their beliefs are bending to the will of the oppressor.
Architecture lives not only as an aesthetic that can be pleasing to the eye but, as a Distant Early Warning system occurring in our civilization to our colony of mitochondria which reflects the collective thinking and experience of a society.
You’ve been warned yet again by me. Are you listening to the whispers of Nature, or will you continue to be seduced by man’s technology?