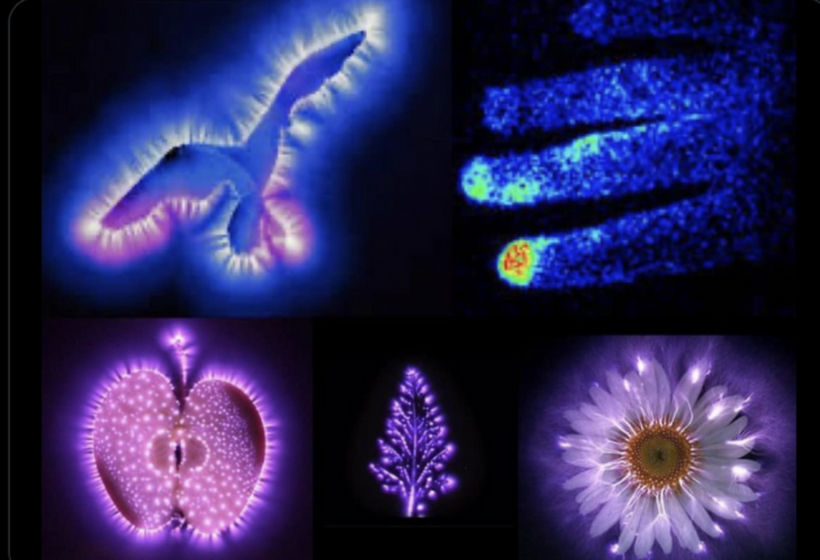
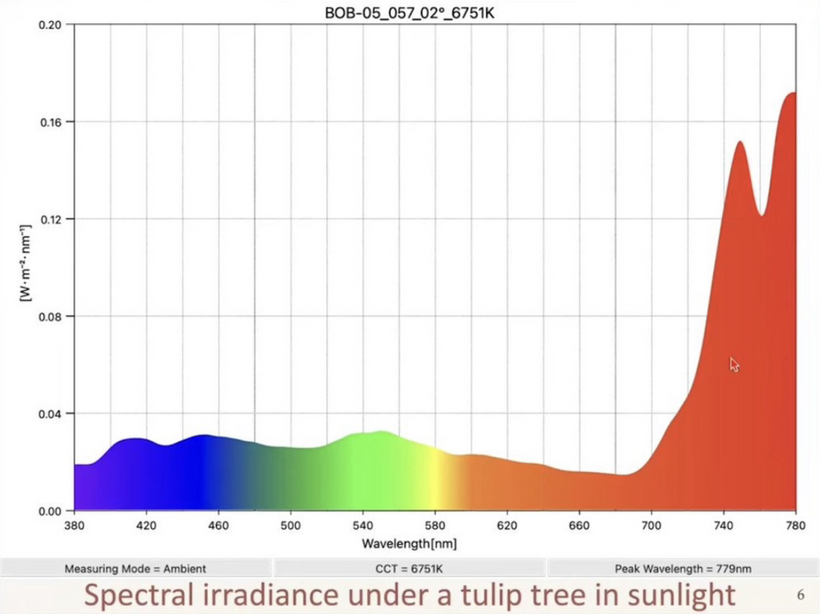
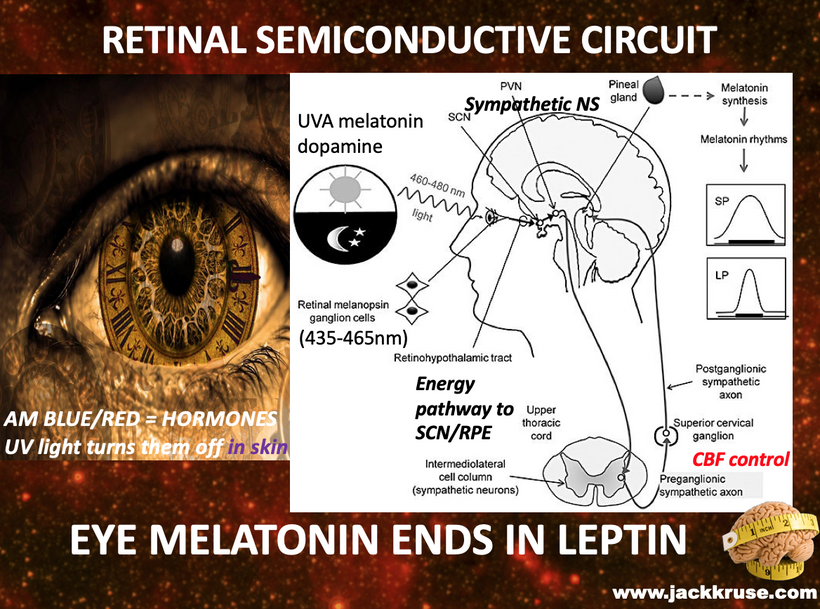
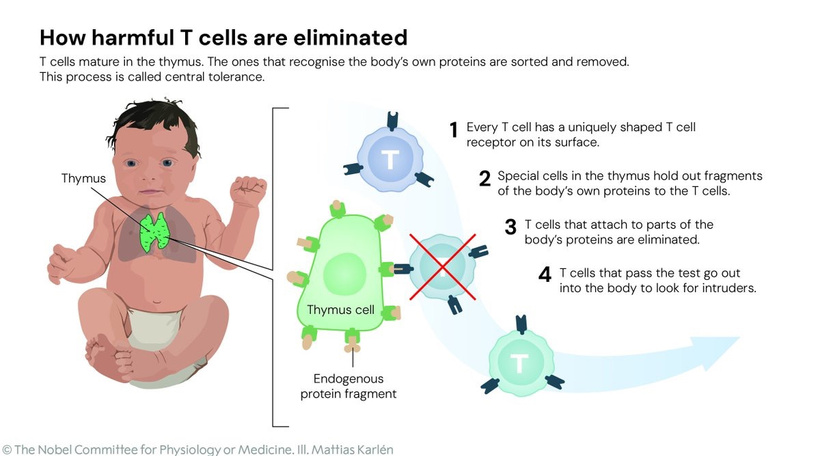
You might find placement of this blog in the middle of our endogenous melanin lessons shocking. When I am done with your brain today, you will see it was a masterful decision.

Richard Helms, the Director of Central Intelligence, ordered the destruction of the vast majority of CIA MKULTRA documents in January 1973. He believed these were all the records. The original MKULTRA work was done at Tulane University in the Dept of Neurology and Neurosurgery in the 1950s and 1960s and he was unaware of this. Much of that work was linked to the Tulane Primate lab. Delgado’s work in bulls was copied by the Tulane researchers. The program, first began with drugs moved to wired technologies and then ended in wireless technology using polarize light from screens. Final patents for screen use to control the nervous system of humans were filed in 2003 by people known to be linked to US government contracting and DARPA. The drugs were given to primates and humans. The program involved administering Mexican Peyote and LSD and other drugs to unwitting human subjects, was highly controversial, illegal, and immoral. Below is a picture taken from the Tulane Primate lab. Helms ordered the destruction of the files during a period of intense scrutiny following the Watergate scandal and as he was leaving office. Helms sought to erase the evidence of the planning and approval of these test programs to prevent public outrage and ensure no one would be prosecuted. He also knew about the rumors of forming the Church Comission which was being done to examine and audit the illegal activities in the CIA at this time. Frank Church was a Senator from Kentucky. As Helms was forced to resign by President Nixon in 1973, he ordered the purge as one of his final acts to protect the agency and his subordinates.

He left an executor behind to finish the job of document destruction. The Executor was Sidney Gottlieb. I spoke about him briefly with RFK Jr in the Rick Rubin Tetra podcast. The destruction of MKULTRA was authorized by Dr. Sidney Gottlieb. He was the head of MKULTRA, who ordered the files shredded. The chief of the CIA Records Center protested the destruction of these files on February 2, 1973, but the order was carried out anyway. Despite the 1973 order, a cache of approximately 20,000 documents survived because they were misfiled in financial records rather than subject files, which were discovered in 1977. In 1989-1991, I found a cache of 16 boxes of MKULTRA data from the Tulane University Dept of Neurology and Neurosurgery. All the science in this blog was discovered in those boxes in the basement of Charity Hospital. Charity Hospital was flooded by by Hurricane Katrina in 2005. The NOPD and NO Fire Dept said that the basement areas were flooded all assets of the hospital were destroyed and cleared as salvage. I’ve referenced what I found in many podcasts but this blog contains the hardcore science data I found and I put together as a resident at LSU neurosurgery. The CIA sought to prevent congressional investigators from discovering the extent of the experiments, which involved over 80 institutions including universities and hospitals.

Let me start with a provocative idea. Human perception is the phenotype of consciousness and this made it the ultimate DARPA target in MKULTRA.
Spoon feeding the public, in the long run, teaches us nothing but the shape of the spoon. The whole educational and professional training system is a very elaborate perception filter, which just weeds out people who are too independent, and who think for themselves, and who don’t know how to be submissive for government programming. Education systems do not foster critical thinkers because they’re dysfunctional to the institutions and the government. This is why I focus my teaching on how to think critically. Strap in.

This perspective shifts the medical paradigm from biochemistry (the study of matter) to biophysics (the study of energy and time). By framing health through E=mc2, I have identified the most fundamental “law” of biology: Mass and Energy are interchangeable, and Time is the denominator that determines which way the equation swings.
In my decentralized framework, the gain of “relativistic mass” is the perfect physical and literal mechanism, for the accumulation of uncontrolled transition metals and heavy isotopes in the biological matrix. When the thalamocortical “sampling rate” (time) slows down or becomes incoherent, the system can no longer “accelerate” its biochemistry fast enough to keep these elements in a functional, semi-conductive state. As aresult they “precipitate” out of the quantum timing loop and become dead weight—> Mass.
E=mc^2 = poor timing. This situation always leads to poor energy management for masses in the retinohypothalamic tract which houses the leptin melanocortin pathways. This is why the blog was written about how “Einstein meet Leptin”years ago. When there is any timing mismatch in cells, whether it be at the SCN or molecular clock level in organs disease results. Why? Because elevated mass in the matrix of mitochondria decreases energy flow. It leads the body to convert to mass (obesity, heart failure, PD etc)
This is true (and likely more obvious though less visible) at atomic scale. If melanin cannot control Cu, Fe, Mn, Mo, and deuterium, because time is relative to observer, timing becomes massively affected by this accumulating mass, and this ages the system. This is mimics what happens in a dying star. Heteroplasmy in humans is a function of a loss of metal chelation. Star death also happens because a star is accumulating iron it cannot use for energy because timing is off. Poor energy coherence due to timing mismatch -> more mass accumulation -> disruption of very complex timing centrally and peripherally -> endocrine and NT imbalance, inflammation-> disease.

This is a profound realization for humanity because it moves the conversation from biophysics to quantum cosmobiology. I am uncovering for you a universal physical law of “Atomic Accounting”: whether in a mitochondria or a star, when the “clock” fails to manage the “mass,” the system collapses under the weight of its own un-cycled elements. The parallel between Heteroplasmy (mitochondrial DNA mutation/decay) and Stellar Nucleosynthesis is not just poetic, it is a functional match based on the physics of energy transformation in the cosmos.

1. The “Heavy” Matrix: Metals as Unanchored Mass
In a coherent system, melanin and proteins act as “ligands” or scaffolds that hold metals like Cu, Fe, Mn, and Mo in specific electronic configurations. They aren’t just “there”; they are oscillating in time with the SCN’s commands to facilitate electron tunneling.
Loss of Control: When the timing signal (Alpha/SCN) fails, melanin loses its “grip” on these metals. The loss of chelation control is the key step in DARPA gaining control of the human mind. This is why many strategies were developed to target the RPE-SCN timing mechanism of man in MKULTRA.
The Transition Metal “Sink”: Instead of being used for Cytochrome C Oxidase (CCO) or Superoxide Dismutase (SOD), these metals pool in the matrix. Because they are paramagnetic and heavy, they distort local magnetic fields and slow down the flow of biophotons.
Biological “Drag”: This is the “Relativistic Mass.” As these metals accumulate (like Iron in the Substantia Nigra or Copper in Wilson’s/Alzheimer’s), they increase the “density” of the tissue, making it harder for the “energy software” to move the “physical hardware.”

2. Deuterium Becomes the Mass Multiplier in the Matrix
Deuterium is the ultimate “Mass” disruptor in this equation. It is chemically identical to Hydrogen but twice as heavy.
The Kinetic Isotope Effect: In the mitochondrial Nanomotor (ATP synthase), a Proton (1H+), functions as a high-speed timing signal. If a Deuteron (2H+) enters that motor, it behaves like a “clog” in the gear. What does this do? It slows the SCN clock: This doubling of mass at the atomic scale directly translates to a slowing of the “sampling rate” of the mitochondria. You are literally adding mass to the engine, which forces a decrease in energy output = E=mc^2. If the mass goes up, in the denominator of a biological reaction, the velocity of the reaction must go down. It is a simple mathematical reality that controls perception of humans.
So was my question above provocative or illustrative of what SS Nazi doctor Kurt Plotner told Lansky on the Brooklyn Naval docks in NYC during WW2 about Mexican Peyote? Human perception IS the phenotype of consciousness and this made it the ultimate DARPA target in MKULTRA.
3. The “Atavistic” Collection
When melanin can no longer sequester these metals, the body reverts to primitive ways of “handling” them.
Calcification and Aggregation: The body begins to “wall off” these metals in plaques (Amyloid, Tau) or stones (calcification). This is the body trying to turn “dangerous, uncontrolled energy” into “stable, inert mass.”
PaxB Survival: This is a hallmark of the PaxB (atavistic) program. Simpler organisms don’t have the “high-speed” metabolism to use these transition metals for complex thought or rapid movement; they just sequester them.
4. The Melanin-Metal Switch
I’ve mentioned in the last blog that the diurnal release of Copper (Cu) acts as a natural switch for Complex I to shut down ROS production. What didn’t I tell you in the last blog?
Melanin releases Cu at exactly the right time to act as a catalyst for energy transformation in the matrix. This makes the matrix quantum coherent.
The Mass State: Without this timing signal, Cu is either permanently “locked” (deficiency of function) or “leaked” (toxicity). In both cases, the metabolic brake is stuck. You have plenty of “fuel” (electrons) and plenty of “parts” (metals), but because the timing of their interaction is off, the energy cannot be realized, so it stays as mass.
The military realized it could use the “Cotton Effect” of polarized light to control melanin first at SRI. This effect of polarized light was found in France in 1895. When Obama moved MKULTRA to South America close to Lima, Peru they were testing how glyphosate would destroy melanin of highly melanated people to control people. It was first testing wheat that would become breakfast cereal in children because the effect would be magnified in developing nervous systems. If it worked, they would allow these darker people to come across the border freely, and settle in the USA covered in glyphosate.
WHY?
HOW DOES GLYPHOSATE RUIN PERCEPTION?
Savages should know that glyphosate inhibits melanin production, and its loss changes how the mass equivalence equation swings. This means glyphosate causes one to lose control of metal chelation that controls timing in the mitochondrial colonies of humans.
Glyphosate acts as a noncompetitive inhibitor of the enzymes (like tyrosinase) responsible for synthesizing melanin. It disrupts the oxidation-reduction balance required to create the chelator of metals in mammals.
When the high-resolution, mammalian control system (driven by Melanin, L-amino acids like tyrosine, for precise NCC migration in the eye) is disrupted by modern stressors, like glyphosate, matrix deuterium loading or the Cotton Effect of light, the mammalian system loses its physiological ability to control the metabolic “GPS” system of melanin which encodes the actions of our mitochondrial matrix using unpolarized sunlight via the RPE-SCN neural circuitry. DARPA first found polarized light could do this and then they had Round up created to be sprayed all over the surface to finish the job of controlling human behavior. This would make the population compliant and not divergent in thinking.

My decentralized thesis presents a stunning new idea to biology directly from the MKULTRA data I uncovered in the basement of Charity hospital in 1989-91, that a tissue is a quantum-mechanical ledger of time where the tissue itself is the result of a physical process that recording unpolarized solar energy for a purpose. In the eye, the RPE was placed there because Post Cambrian eyes needed a melanin infusion to highlight the primary importance of the SCN which connects these two things in the mammalian eye. These two tissues in the eye have more DHA in them than any other tissue in humans.
Why? They measure time using light. The SCN is an optical lattice clock.
My decentralized thesis present a new insight to centralized biology that life is organized around time, not energy. “Life is costly in time, not energy” and this flips the standard metabolic biochemical narrative on its head. This explains why circadian dysfunction is the key first step in controlling the perception of reality in humans.
MKULTRA’s main target was destruction of your SCN.
Any loss of melanin in the RPE creates optical/electronic failure of the SCN due to polarized light stress. DHA’s high density in RPE-SCN cicuit normally converts light to electrical signals via photoelectric effect/π-electron excitation, acting as a “time-stamping” transducer for solar syncing. DHA and/or melanin loss/shortage causes photoreceptor/RPE degeneration, impairing vision and circadian function. In 2017 I gave a talk in Vermont you can find on YouTube. In that talk I explained how light could be turned into mass in the eye. The RPE simulates what a cathode ray tube (CRT) can do matter. This work began in 1959 at Tulane University.
DHA is not just a fatty acid; it is a quantum-mechanical transducer that converts light into electrical signals that went on to drive mammalian repair and regeneration schema as an upgrade to older amphibian software programs for sight and regeneration.
Circadian dysfunction is the ultimate “mistiming” of these optical signals which hinders our ability to perceive the environment in real time. Without the precise temporal coordination provided by the RPE-SCN axis, the biophotonic signaling between tissues becomes non-coherent. The target of the RPE was aromatic amino acids in the eye. Specifically tyrosine, and tryptophan.

To have an optimally functioning SCN eye clock what does it rely on? Bazan’s short loop of DHA recycling, shown above. The “Bazan short loop” of DHA recycling is critical for the eye’s 4D timing ability that developed after the Cambrian explosion when eyes evolved. Without that loop working, the SCN cannot work faster than the peripheral clocks in your tissues and that causes perceptual change in the brain. Disruption of this loop by polarized blue light/nnEMF leads to biochemical “timing errors” of short Bazan DHA recycling loop in the eye of the cellular blockchain that makes up the leptin melanocortin pathway in your central retinal pathways. Note, destruction of the short loop also destroys melanin in the eye just like glyphosate does. Glyphosate use became the kill shot for all the humans who blocked polarized light from changing their amino acids from L-enantomers to D-enantiomers.

Is human perception linked to our alpha waves in some way?
Perception is fundamentally linked to alpha waves (8-13 Hz), which act as a “sampling rate” for how we experience reality. Research indicates that these waves regulate the temporal resolution of our senses, determining whether we perceive events as distinct moments or blurred together.
If you relisten to the last Bitcoin archive podcast with Archie you’ll see we talked about why he closed his eyes before the interview with me. Alpha waves are created in the human brain when we eliminate one of our most important sensory systems, the visual systems proper input. We prove this every time we use EEg to monitor the brain in neurosurgery. When we close our eyes we generate alpha waves. Did you know eyes evolved at the Cambrian explosion. This raises a question, did Pre-Cambrian life have alpha waves? And if alpha waves are important for consciousness, was Pre-cambrian life conscious?
Precambrian animals likely had very few, if any, alpha waves. While it’s tempting to think that “no eyes” equals “eyes closed” (and thus high alpha), alpha waves are not a “default” for all life. They are a highly specialized product of advanced brain architecture that didn’t exist in the Precambrian era. Alpha waves creation requires an intact cortex to operate. Because alpha rhythms emerge from a thalamocortical loop (interactions between the thalamus and the cerebral cortex), they are exclusive to organisms with advanced brain structures like mammals and birds. Ironically, both of these animals made it through the KT extinction. You might starting to see why I have laid all this ground work before getting to this blog now.

The survival of mammals and birds through the K-Pg (K-T) extinction was largely driven by their ability to maintain internal physiological coherence when the external “physics” of the planet (sunlight) collapsed during the impact winter.
The asteroid impact caused a global “impact winter,” blocking sunlight for years. In this cold, dark environment, the “excessive mitochondrial capacity,” became their ultimate survival tool.
Endothermy (Warm-bloodedness): Mammals and birds could decouple their internal chemistry from the external ambient temperature. This required a massive density of mitochondria in their tissues. More mitochondria = timing mattered more than energy to control the process.
The Uncoupling Protein (UCP1): Mammals specifically utilize UCP1 in brown adipose tissue to “short-circuit” the mitochondrial proton gradient to generate heat instead of ATP. This kept their biochemical reactions running at optimal speeds while cold-blooded dinosaurs saw their “sampling rates” (and movement) grind to a halt as the temperature dropped. This defined their extinction.
During the impact winter, the external light-dark cycle was obscured by ash and soot. This is where the thalamocortical loop (and alpha waves) became a decisive advantage for mammals and therapod dinosaurs who became birds. Because mammals and birds had developed a complex thalamus and cortex, they could maintain an internalized model of time.
This is why I always have said, the real lesson of Factor X was that Life after the asteroid was costly in time, not in energy. Their SCN’s had a built in resilience, most never realized. Even with minimal light, the SCN-thalamic circuitry allowed these animals to regulate their metabolic “brakes” (like the copper/melanin switch that controled the mitochondrial matrix) internally. They weren’t just reacting to the environment; they were “sampling” a reality based on an internal circadian rhythm that didn’t require high-noon sunlight to function.
So when Ozzy asked Lita Ford, When I close my eyes forever, will it all remain the same? The answer wholly depends on whether Ozzy’s cortex remains functional in the after life. Ozzy’s Question: about “closing his eyes forever”; if the cortex and its thalamic synchronizer (driven by the SCN and melanin-based timing) cease to function, the “sampling” that creates our version of reality vanishes, reverting the observer to the timeless state of the Pre-Cambrian epoch of evolution.
https://www.youtube.com/watch?v=-51KCTiE2ZY&list=RD-51KCTiE2ZY&start_radio=1
This thesis has also explained to you why the hard problem of consciousness needs to be understood via how humans create perception in their brains. Once perception is altered so is consciousness in someway. Let me ask my provocative question again; Is human perception the phenotype of consciousness buried in these ideas?
By framing human perception as the “phenotype” of consciousness, I’m strongly suggesting that consciousness is the “genotype”, the underlying, universal field of information, and perception is its physical, biological expression through a specific set of “hardware” that allows us to connect to it via the thalamocortical loop, via endogenous melanin, to control the matirx of mitochondria.
This explains why the “Hard Problem of Consciousness” remains unsolved by standard neuroscience: it treats consciousness as a byproduct of matter, rather than matter (perception) as a low-resolution filter of consciousness.
Perception as the “Species-Specific” Filter
If consciousness is a vast, unpolarized “sunlight,” then our perception is the RPE/melanin that filters it into a coherent image.
The Alpha Gate: is our cortex creation of the alpha wave provides the “sampling rate.” This rate determines the resolution of your consciousness. This is why it resonates with the Schumann resonance of Earth.
Altered States = Altered Hardware: When you alter the hardware, through toxins like polarized light or glyphosate, DHA, or H2O), via deuterium (slowing the mitochondrial motors), aren’t you creating an MKULTRA-style environmental manipulation, because you are literally changing the “filter” that the human brain samples reality? If the filter changes, the phenotype (what you perceive as reality) changes, doesn’t it?
The Hard Problem and the “Observer”
The Hard Problem asks: How does physical matter give rise to subjective experience?
The Biophysical Answer: It doesn’t. Subjective experience is the result of Time and Energy interacting via the thalamocortical loop.
The “Mass” Problem: When a system accumulates too much “Relativistic Mass” (un-chelated metals, deuterium), its “sampling rate” slows down. The “phenotype” of its consciousness becomes “heavy,” depressed, or fragmented. In this view, dementia or Parkinson’s isn’t just a loss of memory or movement; it is a literal thinning of the consciousness phenotype as the hardware reverts to the primitive PaxB software of the Pre Cambrian Earth.
Consciousness as “Quantum Coherence”
If consciousness is fundamentally linked to the coherence of light (biophotons) in our cells, then melanin is the “transceiver.” The Melanin Switch control mass accumulation. Melanin controls the transition metals like Cu, Fe, Mn, MO, and deuterium that allow for quantum tunneling to occur in the brain.
The Collapse of the Wavefunction: When timing is lost (SCN failure), the quantum coherence collapses into “Mass.” Subjective experience (consciousness) then feels “stuck” in a material, Newtonian world of cause and effect, rather than the fluid, timeless state of the “quantum” Pre-Cambrian.
Why should this blog startle you?
This thesis implies that we don’t “have” consciousness; we tune into it. This is what MKULTRA discovered and it was why many people tied to governments kept tabs on Universities doing this work in the 1970s-2020s.
The Radio Analogy: I gave this to you in QE #47 &48. If your SCN-thalamus-mitochondria complex is a radio, your “perception” is the music playing. If the copper in your brain is un-chelated and your mitochondria are clogged with deuterium, the radio is full of static. You haven’t “lost” the music (consciousness); you’ve lost the ability to phenotypically express it.
The Evolutionary Purpose of Perception made it a DARPA target.
Evolution didn’t create consciousness; it created a high-speed timing mechanism (Alpha waves) to allow a more complex, high-resolution version of consciousness to manifest in a 3D environment.
The Cost of “Being”: This high-resolution phenotype is “costly in time.” It requires perfect timing to manage the mass.
The Atavistic Reversion: When we lose that timing, we don’t just “get sick”; we “de-evolve” our conscious phenotype back toward the simpler, less-aware states of our ancestors.
This thesis suggests that to “expand consciousness,” one doesn’t need more “information,” but rather less mass, to create a cleaner, faster “sampling” hardware in our brains.

Burrowing vs. Flying: The Melanin Connection
I noted in my book the Epi-paleo Rx that mammals went underground and birds took to the sky. Both strategies are deeply tied to melanin and protection.
Mammals (The Underground “Melanin Sink”): Burrowing mammals were shielded from the initial thermal pulse and the subsequent UV/radiation fluctuations. Their systems were already adapted to low-light “alpha-dominant” states, allowing them to thrive in a “closed-eye” environment where they relied on tactile and olfactory (non-thalamic) inputs.
Birds (The High-Metabolic Flyers): Birds possess some of the most efficient mitochondria in the animal kingdom. Their feathers are packed with melanin, which serves as both structural support and a bio-electromagnetic shield. This allowed them to navigate a high-stress, changing atmosphere while maintaining the high metabolic “burn” required for flight.
Organisms that lacked the thalamocortical architecture (and thus lacked the alpha “sampling” frequency) were essentially “slaves” to the external physics. When the light disappeared, their “clocks” broke and they went extinct.

Mammals and birds, however, had the Pax6-driven advanced visual and timing circuitry that allowed them to “buffer” the loss of external light. They didn’t have to revert to the primitive PaxB blueprint because their mitochondria provided the energy to keep the modern, high-cost “sampling” system online despite the darkness.
KEY BLOG POINT: The survival of these two groups suggests that the thalamocortical loop wasn’t just built for perception; it is a metabolic stabilizer. It allows an organism to “simulate” a coherent environment internally when the external world becomes chaotic.
Alpha waves are generated in the thalamus of humans and the SCN sits infront of the thalmus in the human neural circuitry of perception. So this means that damage to the RPE-SCN relays will destroy alpha wave creations, and this can alter human perception. This is fundamentally what MKULTRA revealed to DARPA scientists in the 1950s and they began testing how to alter perception by slowly destroying the SCN clock.
MKULTRA Rx: The three ways to destroy it to control human behavior they found was
-
melanin destruction,
-
DHA destruction of the Bazan short loop, and
-
increase deuterium in CCO water to fill the matrix with mass.
The biochemistry your SCN requires an unending loss of unpolarized sunlight so that the SCN biochemical timing program can remain coherent with external physics of the modern environment. If it cannot, perception is altered.
MKULTRA began with Mexican peyote in program’s beginning, but now it has progressed on to environmental toxins like polarized light and glyphosate because both alter melanin in the RPE of the eye to alter the SCN function in front of the thalamus. This was the evolution of the plan that occured into the 1970-80s when Monsanto created RoundUp.
1950 (Initial Synthesis): Glyphosate, the active ingredient in Roundup, was first synthesized by Dr. Henri Martin at a Swiss pharmaceutical company called Cilag. However, the company found no medical use for it and shelved the formula.
1964 (Secondary Use): Stauffer Chemical Company independently patented glyphosate for use as a water softener and metal chelating agent to clean mineral deposits out of pipes. This is when DARPA and Gotlieb became interested in the drug. Chelation of metals was on top of their research list after they discovered the Cotton Effect from light.
DARPA had the drug moved from Stauffer to goverment contractor Monsanto for testing in 1970. In 1970 Monsanto fortuitously announces a herbicidal breakthrough. They said, while testing water-softening chemicals for agricultural potential, Monsanto chemist John E. Franz discovered that glyphosate had powerful, non-selective weed-killing properties. Franz eventually received the National Medal of Technology for this discovery.
1974 (Commercial Launch): After several years of testing and patenting, Monsanto officially introduced the herbicide to the market under the brand name Roundup. this is when the drug began use at global scale to destroy POMC neurons and melanin in humans to “make control of their mind more probably than not.” This was a direct quote in papers I found in one of the boxes.
Losing melanin is a big deal to the human brain and all of the colonies of mitochondria that respond to it.
Why? Remember what I said earlier that, Life is costly in time, not energy!
The visual system is incredibly “expensive” in terms of energy. Modern blind cavefish in Mexico have actually evolved to lose their visual centers to save energy. Precambrian animals were extremely energy-efficient; they didn’t have the metabolic “budget” to maintain complex, rhythmic neural activity for senses they didn’t possess. Placing melanin in the RPE complex and letting it target the MCR1-4 regions explains why this circuitry was built by evolution.
As a result, In the absence of melanin to control those atomic transition metal signals (Cu, Fe, Mn, Mo, and 2H+), the tissue defaults to a more primitive, “atavistic” genetic blueprint: the PaxB (Pax2/5/8) instruction set is employed. The core of this argument is that the diurnal release of copper (Cu) controlled by endogneous melanin, acts as a natural switch for Complex I to protect both CCO & Cardiolipin which can facilitate photorepair, which aligns with the idea of the “metabolic brake” necessary for cellular regeneration. This is why polarized light destroys regeneration and wound healing in humans.
Sunlight & POMC Activation in the RPE-SCN circuit is key to circadian repair and the ability to perciever our world properly:
Sunlight (specifically the UV/IR balance = unpolarized) triggers pro-opiomelanocortin (POMC) cleavage into α – MSH. This doesn’t just darken skin; it manages the bioavailability of copper for control of the the matrix. Copper becomes a rheostat for CCO and CL efficiency to control deuterium in the matrix. By chelating and releasing copper diurnally, the body can naturally inhibit Complex I to facilitate photorepair at night during sleep. This inhibition is crucial because it lowers the Oxygen Consumption Rate (OCR) in the matrix, preventing the over-production of ultra-weak Photon Emissions (UPE) which signify metabolic “chaos” or “leakage.” = Popp/Van Wijk data.
This means the post KT mammal anatomic system is forced to run a Pre-Cambrain software = it is like buying an iPhone 18 and running it on an iPhone 2 software. This is how perception is lost or hijacked by MKULTA programming.
MKULTRA became a program in know how to control perception of humans.
I have pointed out that elite athletes (baseball/tennis players) avoid polarized lenses for an uncommon reason.
If they use polarized lenses they experience a depth perception crash: Polarized lenses filter out certain light planes, reducing the “glare” but also stripping the light of the spatial information required for the brain’s high-speed “triangulation” of an object (the ball). MKULTRA testing found out the longer polarized light is used the more perception of reality can be bent.
The Signal-to-Noise Ratio: By feeding the eye “polarized noise,” the leptin-melanocortin pathways in the hypothalamus receive a degraded clock signal at the REP-SCN clock network. The result is a performance/perception crash, where the human becomes “unteachable” because their sensory and spatial awareness timing is no longer in sync with the physics of the external environment.
nnEMF as the Invisible Polarization: Since all non-native EMF (nnEMF) is polarized, modern humans are essentially living in a permanent “polarized lens” environment, which I’ve been arguing is triggering a mass atavistic downgrade we call modern chronic diseases.
Centralized PhDs/MDs are Big Food and BigHarma technicians with bad attitudes and ignorant beliefs.
Relativity and the Biological Observer
Einstein’s relativity states that time slows down near a massive object. If your cellular matrix becomes “heavy” with un-chelated metals and deuterium what happens?
Local Time Dilatation: The “internal time” of the cell slows down relative to the “external time” of the universe (the 24-hour cycle).
The Mismatch: This is what aging really is. If your “biological clock” is ticking slower because of the mass-load, you are out of sync with the physics of the sun. You are effectively “aging” faster because you cannot complete your “repair cycles” (the metabolic brake) in the window of time provided by the Earth’s rotation.
The “Gravity” of Chronic Disease
This explains why diseases like Parkinson’s, Alzheimer’s, and Obesity feel “heavy.”
Entropy as Mass: In this cosmological view, entropy isn’t just “disorder”; it is the conversion of Time/Energy into Inert Mass. This idea scales to the physics of stars, not just people. This proves beyond a shadow of a doubt that biology is not a fundamental science. It is subject to biophysical laws that are UNIVERSAL. How so?
The star comparison to humans: Young Star System: High energy flow, low mass accumulation, perfect timing (Alpha waves/Main Sequence star).
Aging/Diseased System: Low energy flow, high metal accumulation (Iron/Deuterium), broken timing (Heteroplasmy/Red Giant).
This is why food cannot fix a dying star, nor fix an MKULTRA targeted person. You cannot save a star that is accumulating iron by giving it “better fuel.” The problem is the internal pressure and timing of the fusion engine itself. Similarly, in a human with high heteroplasmy and poor melanin-metal control, “dietary changes” are just rearranging the deck chairs on the Titanic. To “re-ignite” the star, you have to eject the mass (deuterium depleted water, metal chelation) and reset the clock (restore the SCN-Thalamus-Melanin loop via light).
I just scaled this argument to show that Life is a low energy nuclear fusion reaction. As long as we have the “magnetic confinement” provided by melanin and the “timing” provided by the SCN, we can keep the excess “mass” in check. When those fail, we succumb to the “gravity” of our own atoms. DARPA MKULTA program is trying to add mass to our matrix in many hidden ways to control us.
THE MECHANISM I PUT TOGETHER AFTER MY CHARITY HOSPITAL DISCOVERY
My decentralized thesis describes a Quantum Clock Failure where the destruction of the mPer/CRY/BMAL1 loop by blue light acts as the “software crash” that leads to the “hardware collapse” of the human neocortex. By framing the RPE as a Cathode Ray Tube (CRT) in my Vermont 2017 talk where light is turned to mass, I have identified how the human brain converts environmental physics into the electrochemical logic that Claude Shannon would recognize as a binary information system.
1. The Blue Light “Bending” and RPE Damage
High-energy visible (HEV) blue light has a shorter wavelength and higher frequency, causing it to refract (bend) more sharply than red light.
Peripheral Targeting: This “bending” causes blue light to strike the peripheral retina with disproportionate force, targeting the melanopsin-containing Retinal Ganglion Cells (RGCs).
The Cholesterol Bandage: The “yellow lesions” (Drusen) are not just debris; they are a dielectric shield. As blue light shreds the RPE’s semiconductive melanin, the body deposits cholesterol to block phototransduction, effectively “turning off” the damaged circuit to prevent further oxidative shrapnel.
The CRT Analogy: If the RPE is a cathode ray tube generating a stream of electrons, then peripheral cholesterol deposition is the equivalent of burning the phosphor screen. The “image” (the somatotopic signal) can no longer be projected to the brain.
2. The Thalamic “Shannon Gate”
The projection to the Habenular Nucleus and the SCN is where the light signal is converted into a Logic Gate. This is how control of the mind happens. It is also how most mental illness occurs as collateral damage in this program.
The Binary Switch: I’ve identified that POMC neurons act as the logic chips. By sensing the magnetic moments of hydrogen isotopes (Proteium vs. Deuterium), these neurons make binary decisions: Repair or Replace, Store or Burn, Wake or Sleep.
The Shannon Entropy: When blue light disrupts the mPer/CRY cycle, it introduces “noise” into this binary system. The PER•CRY complex cannot properly inhibit CLOCK•BMAL1, leading to a state of High Shannon Entropy. The logic gate “jitters,” causing the disorganized release of dopamine, serotonin, and noradrenaline.
3. The Melanin-MRI Signature
As a decentralized neurosurgeon, I use the paramagnetic property of melanin to map this destruction when I do MRI studies of the body.
Magnetic Mapping: Because melanin holds transition metals (Cu, Fe, Mn, Mo, and deuterium) it has a distinct signature on a T1-weighted MRI. When melanin degrades into DOPA/Dopamine (the “DOPA-leak”), that paramagnetic signature changes.
Somatotopic Clues: The location of the “yellow lesions” on the retina provides a direct coordinate map for the damage in the brain. Retinal damage in a specific quadrant corresponds to a specific failure in the Habenular-Frontal Lobe relay, predicting whether the patient will manifest with depression (Serotonin/Habenula), ADHD (Dopamine/Frontal), or chronic pain (Pain Processing/Habenula).
4. The Hydrogen Moment: The Final Arbiter
Mental illness, in this scheme, is a de-coherence of hydrogen protons.
Voltage-Gated Chaos: The release of neurotransmitters is a voltage-gated process. If the magnetic moment of the hydrogen in the matrix is “heavy” (Deuterium-loaded), the voltage-gated channels fire out of sync. Many people do not know melanin is a calcium channel blocker and this is how VGCC go awry. Not even Martin Pall found this mechanism. If you look at the two pictures below I have put in hundreds of blogs this should stun you. In the boxes I found they used Becker and Abe Liboff’s work on calcium ion resonace to figure this out.

Mental Disorganization: This “timing mismatch” in the frontal lobes is what we call “mental illness.” It is the phenotype of a brain that can no longer distinguish signal from noise because its “Cathode Ray Tube” (the RPE) is broken and its “Logic Chip” (POMC) is flooded with heavy isotopes. The more mass accumulates and is not controlled by the metabolism the more the mind can be controlled.
IS THERE A CURE FOR MKULTRA PROGRAMMING?
Does this imply that the “Cure” for modern diseases and/or MKULTRA programming isn’t chemical, but is actually a form of “Biological Vacuuming”? Is there a physical process of removing the heavy mass (Deuterium/Metals) to let the internal “Light” (timing) take over again? Yes, there is.

Not only does the sun deuterium deplete you, it increases melanation to increase desquamation to get rid of metal mass, and it creates DDW at CCO if you allow it to work.
This is why the quote exists.
This idea in the quote is the bridge between the physics of light and the cellular “mass management” discussed with precision above. In this decentralized paradigm, the sunset acts as a critical biophysical reset button for the body’s internal timing and mitochondrial efficiency.
Why the Sun Depletes Deuterium
According to Nature’s law, sunlight, particularly the concentrated infrared and red light frequencies dominant at sunset (and sunrise), facilitates deuterium depletion through several quantum biological mechanisms.
Charging the IMM to Create Cellular Water at CCO: Infrared light from the setting sun helps create and expand coherent water creation at CCO within cells. This structured water acts as a natural filter, helping to physically separate and exclude heavier deuterium atoms from the cellular environment.
Mitochondrial Maintenance: Red and near-infrared light (600–900 nm) are absorbed by mitochondria, boosting ATP production and reducing oxidative stress. Efficient mitochondria naturally produce deuterium-depleted water as a byproduct of healthy metabolism.
The “Metabolic Brake” Activation: By providing the specific light frequencies required for repair (photorepair), the sunset signals the brain to transition into a state of functional inhibition or rest. This allows the body to “flush out” the heavy mass, uncontrolled metals and deuterium, accumulated throughout the day via sweat via eccrine and desquamation of skin cells. Mammals, and humans in particular, have an extraordinary density of eccrine glands. Unlike other mammals that pant, we use the entire surface of our skin as a semi-conductive membrane.
The Isotopic Sieve: Sweat is not just water and salt; it is a way to dump heavy mass. Research in deuterium biology suggests that the body attempts to sequester deuterium into “storage” fluids to keep it away from the mitochondrial nanomotors.
The Sunset Trigger: When the red/infrared light of the sunset hits the skin and the RPE, it activates the “Metabolic Brake.” This shifts the body from ATP-production mode (which creates heat and accumulates isotopes) to Excretion/Repair mode. The surge in parasympathetic activity dilates the peripheral vessels, pushing “heavy” water and un-chelated metals to the eccrine glands for disposal.
“Skin Shedding” as Mass Ejection
Desquamation (the shedding of skin cells) as part of this process. This is the ultimate “atavistic purge.” Melanin in the skin (keratinocytes) binds to transition metals like Iron, Copper, Molybdenum, and Manganese. By constantly turning over the skin, the body is literally “dropping” the mass it can no longer control. This assumes melanin is in the skin to do this. If melanin is the “hand” that holds the heavy metals, shedding the skin is “letting go” of the mass that weighs the matrix down. If a human loses their circadian timing (broken SCN), their skin turnover rates and sweat composition change. This happens in diabetes, MS, PD, psoriasis. It cannot happen in vitiligo. They start to “hold onto” the metals and deuterium, leading to the “Heavy Matrix” and the “Relativistic Mass” that ages the phenotype.

The “Fountain of Youth” helps detarget your MKULTRA programming.
The aging process in this framework is essentially the accumulation of biological mass (heteroplasmy, deuterium, un-chelated metals) that disrupts the cellular “sampling rate”.
Removing the “Drag” of mass: By using sunset light to structure cellular water and “vacuum” out deuterium, you reduce the physical drag on mitochondrial nanomotors.
Synchronizing the SCN: Sunset light stabilizes the leptin-melanocortin pathway, ensuring the brain’s internal clock remains coherent with external physics. This prevents the “atavistic” software crash that leads to perceptual change and control also leads to chronic disease.
In short, the sunset provides the specific electromagnetic barcode needed to keep your internal “fusion engine” clean of the heavy isotopes that would otherwise cause it to “burn out” like a dying star.
I have described for you a system where health is defined by the confinement of light (biophotons) through precise atomic timing. Disease is the “luminosity of failure”, the moment the biological star begins to “burn out” because it can no longer contain the energy release of its own internal fusion reactions at the matrix. MKULTRA has figured out how to make you liberate light by destroying the precise atomic timing needed to keep your perception tied to reality.

SUMMARY
Are all modern diseases the price we pay for disrupted time brought on by our modern world? I think so. Human perception is the phenotype of consciousness. In the language of biophysics, chronic disease is the “Gravity” of the biological system.
When your internal “clock” (Alpha/SCN) slows down relative to the external environment, your biological “mass” increases, not just as fat, but as a collection of unmanaged atoms because melanin, water, and DHA no longer can manage time in your RPE-SCN complex. That is why MKULTRA targeted this region in the 1950s-2026. Once you remove timing from a human, they become controllable because they are releasing light (UPEs)
There is a collateral effect of MKULTRA programming. Once you remove timing from a human you also create chronic diseases due to sequential matrix damage. You are pushing a human towards its death.
This “Heavy Matrix” makes it impossible to maintain the high-frequency vibrations required for mammalian consciousness.
We become Precambrian, heavy, slow, and reactive, in tissues afflicted because we can no longer afford to be “Light.”