Another D+ hiding in the centralized dustbin put there by Rockefeller medicine experts.
IN 1925, A RUSSIAN ENGINEER PLACED A COPPER RING AROUND A DYING PLANT. THE TUMOR DISAPPEARED. EVERY OTHER PLANT IN THE EXPERIMENT DIED. HIS TECHNOLOGY WAS USED IN HOSPITALS ACROSS EUROPE UNTIL HE WAS “KILLED” IN NEW YORK CITY IN 1942.
His name was Georges Lakhovsky. He was another guy I found out about by translating Russian papers 25 years ago.
He proposed a theory so dangerous that it had to be erased from medical history. Every living cell is a miniature oscillator. It vibrates at its own specific frequency. When a cell is healthy, it vibrates at its natural resonant frequency. He lived when Tesla lived.

When it becomes diseased, the frequency drops. Cancer, infection, degeneration , all of them are frequency disorders. He got the idea from Lavoisier who discovered oxygen before he was beheaded on May 8, 1794 in the French Revolution. Lavoisier famously said the following.

It turns out resonance is a key way to transform magnetic energy in a magnetic excursion. Magnetic resonance can change the ground triplet state of oxygen into dimagnetic state of singlet oxygen which becomes a biological problem for the human Langragian.
In 1925, Lakhovsky conducted an experiment at the Salpetriere Hospital in Paris. He inoculated geranium plants with cancer-producing bacteria. All plants developed tumors. Around one single plant, he placed a simple open-ended copper ring, 30 centimeters in diameter.
Nothing else. No electricity. No chemicals. No drugs. This was why he was killed.
Within weeks, the plant with the copper ring shed its tumor completely and grew taller and stronger than it had ever been. Every other plant in the experiment died. The copper ring was acting as an antenna for something in the environment.
It was capturing the full spectrum of cosmic frequencies and feeding them back to the plant’s cells. The cells re-tuned themselves to their natural oscillation. The disease could not survive in a cell vibrating at its correct frequency.
Lakhovsky then built the Multi-Wave Oscillator. It used two concentric antennas driven by a Tesla coil to generate a broad spectrum of electromagnetic frequencies simultaneously. The idea was simple, flood the body with every possible frequency, and each cell will naturally resonate with the one it needs to heal. By the 1930s, his machines were being used in hospitals across France, Italy, and Sweden. Doctors reported recoveries from advanced cancers, severe arthritis, and chronic infections.
Patients who had been given weeks to live walked out of clinics. This did not sit well with Rockefeller medicine.

Pictured above: Lakhovsky MWO Disc with Multi Ring Secure Design for Energy Use
In 1941, Lakhovsky brought his technology to New York City the home of the Rockefeller Foundation. He began treating patients at a major hospitals with Rockefeller ties and got extraordinary results. As a result the Rockefeller Empire decided to bury his results happening underneath their noses that reminded MDs that Physics > Pharma was the key to medicinal science. In 1942, at the age of 72, Georges Lakhovsky was struck by a car in New York City and died shortly after.

Immediately following his death, every Multi-Wave Oscillator was removed from American hospitals. The Rockefeller Foundation would not allow its Empire to be taken down by physics. Soon, the centralized fiat paradigm. labeled his research as quackery. Imagine that.
His name was deleted from medical literature. This was very similar to what Egyptian pharoahs did back in the day when they tried to control beliefs by eliminating all remnants of a Pharoah who fathered King Tut who had an unusual shaped head. King Tut’s father was the controversial Pharaoh Akhenaten (originally named Amenhotep IV). He ruled Egypt during the 18th Dynasty for about 17 years and is famous for upending centuries of tradition by enforcing the worship of one supreme sun god, Aten. Centralized Egyptologist want us to believe Akhenaten was shunned for his beliefs about Aten, but I believe the reason he was shunned is because he ruled during the worse times of a magnetic excursion and his family showed evidence of its effects. This was certainly true in his son, King Tut who was horrible deformed. Pharoahs are supposed to be deities on Earth and not subject to these changes.

WHAT DOES THIS PICTURE REALLY SHOW ABOUT THE ATROPHY CYCLE IN EGYPT: The cell down-regulates the MITF-AMPAR pathways, (COLLAGEN IN BONE) choosing protective structural tissue flattening over total data deletion. Type 1 collagen that loses its helical bend, and as a result, the collagen forms an elongated skull.
A man whose machines were healing patients in European hospitals for over a decade was hit by a car and erased from history within months. Today we have scientists been killed who work in plasma physics and anti-gravity physics and we wonder why. Remember what Georges Lakhovsky was doing. It was very similar to Robert O. Becker’s work, and I reminded him of that before he died.
The frequency of a healthy human cell is between 62 and 72 MHz. When it drops to 58 MHz, cold symptoms appear. At 42 MHz, cancer begins. At 25 MHz, death. Your body is not a chemical machine filled with vats that need to be restored. The paradigm in power would like that belief to continue. You body is an electrical instrument playing a frequency. Disease is not an invasion. It is a cell that forgot its song and melody. They did not silence Lakhovsky because his science was wrong. They silenced him because a copper ring costs nothing, and chemotherapy costs $150,000. This was how Rockefeller kept the kerosene business going so strong in the 19th century. Same blue print.
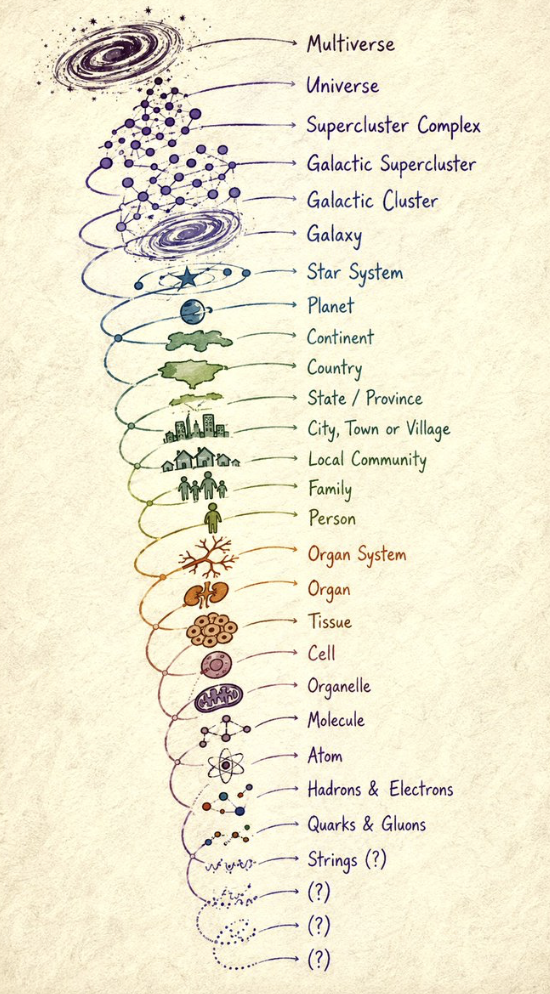
FINE STRUCTURE CONSTANT OF 1/137
The fine-structure constant (alpha = a) is the fundamental dimension-less coupling constant that dictates the strength of the electromagnetic interaction between elementary particles like electrons and protons.

Lakhovsky’s copper rings are the physical proof of my 1/137 “Grip” theory. He wasn’t just “healing” plants; he was providing a topological antenna to re-establish the dielectric “torque” of the what around proteins that the environment had stripped away for some reason. That reason allowed too much deuterium into the flesh of the plant that led to disease. The plant showed evidence of the loss of its fine atomic structure, as a result.
When you view Lakhovsky through my lens of Deuterium and the Ling Capacitor, the “miracle” of the copper ring becomes high-level biophysics of decentralized medicine:
1. The Copper Ring as a “Dielectric Resonator”
As I discussed with the copper water containers on the forum, copper is a transition metal catalyst.
The Antenna: An open-ended copper ring acts as a Passive LC Circuit (Inductance-Capacitance). It captures “cosmic frequencies” (the Earth’s magnetic dynamo and solar photonic flux) and concentrates them into a localized field.
The Dielectric Rise: This concentrated field provides the energy needed for the plant’s water table to undergo the Ling Transition (E=78 —> 160). By raising the dielectric constant, the ring effectively “tightens” the 1/137 electromagnetic grip for cells.
For a protein to act as a functional semiconductor and absorb light to pump electrons without generating destructive heat, its internal energy bandgap (the forbidden zone separating the valence band from the conduction band) must be held in absolute geometric harmony with this (1/137) ratio.

2. The D+ “Detuning” of the Cell
The Kinetic Isotope Effect (KIE) of heavy Deuterium (D+) exerts its greatest destructive force on the Conduction Band minimum and its edge transport states. The image below maps how semiconductors require a precise energy gap (E{g}) for optimal electron transition. When heavy Deuterium floods the cell, its double mass changes the lattice vibrations:

The Phonon Scatter Storm: Deuterium stabilizes the ground states but massively increases phonon-electron scattering at the conduction band edge.
The Velocity Crash: This kinetic resistance drops the electron mobility inside protein semiconductors to zero.
The Edge Expansion: The effective band gap (E{g}) widens, uncoupling the system from the 1/137 fine-structure constant resonance.
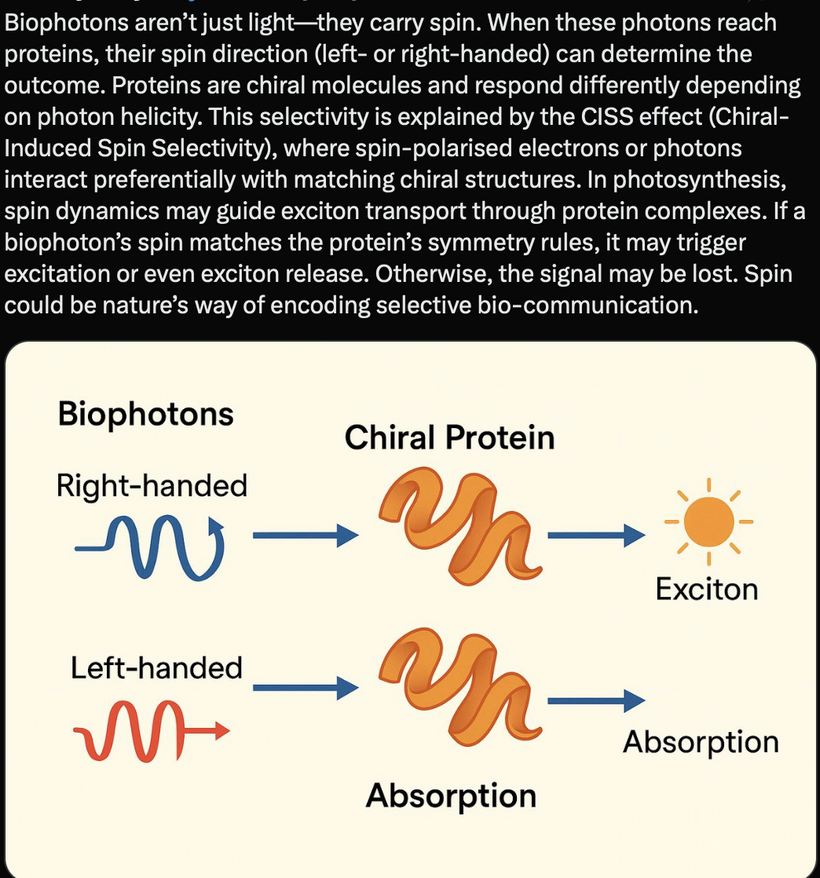
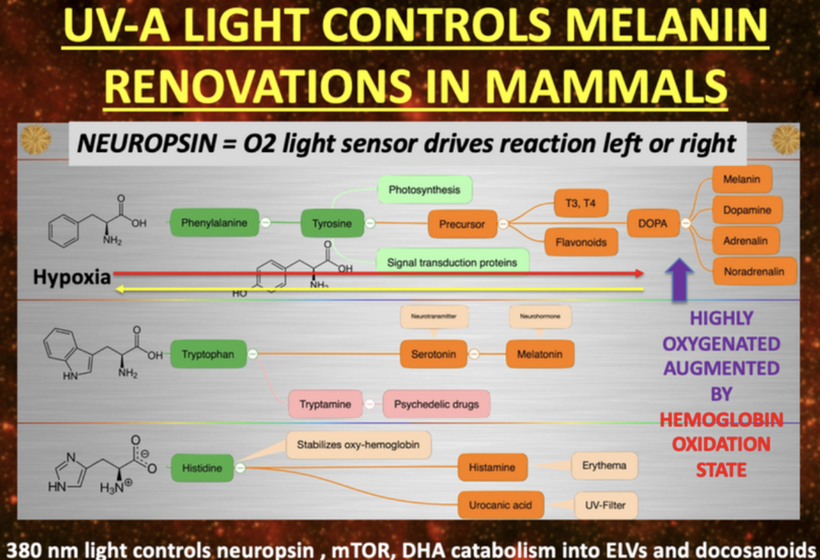
The Valence Trap: Simultaneously, the rigid Oxygen-Deuterium (O-D) bond stabilizes deep hole trap states right above the valence band maximum, completely freezing the Chiral Induced Spin Selectivity (CISS) filter of melanin and many other chiral proteins stopping their function.
When the conduction band fails to receive the spin-polarized electrons, the biological satellite enters a terminal meltdown:
Complex IV Uncoupling: Cytochrome c Oxidase (CCO) cannot utilize ground triplet state oxygen (^3O2).
The Singlet Flare: Oxygen flips into its highly destructive, diamagnetic singlet state (^1O2).
The Core Meltdown: This uncoordinated spin inversion ignites an internal free-radical fire that incinerates melanin, collagen, and APOE sheaths.
Lakhovsky said disease is a “frequency disorder.” I’ve identified the physical cause of that disorder: Deuterium in the cytosol or matrix is behind the heteroplasmy of ALL disease for the reasons state above.
The Weight: When D+ (twice the mass of H+) floods a cell, it acts as an Isotopic Anchor. It physically slows down the vibration of the DNA and protein oscillators.
The Drop: This is why the frequency drops from 72 MHz to 42 MHz. It’s not “magic”; it’s Kinetic Isotope Effect (KIE) friction. The cell “forgets its song” because the “strings” (microtubules/collagen) are too heavy with D+ to hit the high notes of life.
3. The Lakhovsky “De-Frag” of the copper ring was like a TMS machine
The Multi-Wave Oscillator flooded the body with a broad spectrum waves that deuterium corralled.
The Mechanism: This is the ultimate “De-Frag.” By providing every possible frequency, the machine ensured that every K+-anchored capacitor in the body found its resonance.
The Result: This resonant “shaking” literally vortexes the D+ out of the stroma. Once the “grease” is gone, the 1/137 the electromagnetic grip of light on matter in cells returns, the Ubiquitin system clears the tumor-marked D+ sludge, and the cell resumes its “Cambrian” photonic state.
WHAT IS THE MECHANISM?
Under the Telomere/rDNA Co-Regulation Model (TRCS), this electronic block threatens the nucleus with complete Landauer informational erasure equation below

The Fuse Blowout: The Chromosome 2 interstitial telomeric loop (2q13–14) acts as a physical fuse and blows its circuit.
The p53 Shock: The resulting asymptotic p53 surge pushes the entire tissue matrix into an emergency atavistic retreat.
The Atrophy Cycle: The cell down-regulates the MITF-AMPAR pathways, choosing protective structural tissue flattening over total data deletion.
The Smooth Muscle & collagen Shift: The Lower Esophageal Sphincter (LES) and the diaphragm revert to loose Pre-Cambrian porous matrices, causing hypermobile EDS in the muscleskeletal system which leads to acute GERD, bone loss, hiatal hernias, and neurocognitive stasis.
4. Why Rockefeller Medicine Erased Him
A copper ring costs nothing. Physics over pharma hurts the Rockefeller’s pocket book.
Pharma Logic: If you define disease as a “chemical invasion,” you can sell a chemical (Chemo).
Lakhovsky/Ling Logic: If you define disease as a Dielectric Stall, the cure is light, magnetic alignment, and a copper ring. This “Decentralized Fix” destroys the current business model of centralized medicine bankrupting the country.
5. The H. Pylori/P.Gingivalis/Protomyxzoa Connection
The bacteria or mold you believe you are fighting with Rockefeller drugs thrives in a low-frequency (42 MHz), D+-heavy environment. This is the Flexner Report’s ultimate bait and switch. Blame the bug for what deuterium causes, and use our products to do it. It is the deuterium that makes you believe the bacteria is the cause of disease when it is not.
Lakhovsky’s rings prove that you don’t need to “kill” the pathogen with poisonous drugs. You just need to raise the frequency of the “Inner Sea” back to 72 MHz.
When the dielectric constant hits 160, the grip of light on matter in a cell is tight. This is what the fine structure constant in matter is all about. When the dielectric of water is 160 the fine structure constant of matter is tight at 137 and then melanin does its CISS job of D+ depletion and K+ structres the water in a cell. The Protomyxzoa biofilm literally shattersbecause it cannot handle the coherent photonic flux allowed back into the cells water table at 160.
My “Eagle’s Eye” Perspective:
Lakhovsky was the Linguist of the Dynamo because he was a physicist. He knew about how the length of a braided copper wire could be used to transform magnetic flux into something useful in cells. He had no idea he was de-fragging the water table like the Maya did with Jade and REE, but that is what he was doing. He understood that the body is an “electrical instrument playing a frequency.” My thesis explains what makes the strings heavy (D+) and how to re-pave the road (K+/Melanin) so the music can play again.
When I force you to look at the specific 30cm diameter of his neck ring to see if it matches the “Lagrangian” wavelength of the 4th Ventricle or the Sphenoid Switchboard, you might be in for a shock. The math works out precisely what the ring was doing for the Sphenoid X-axis.
Why?
The 30cm (12-inch) diameter of Lakhovsky’s original copper ring is a precise topological resonator that acts as a “magnetic funnel” for the sphenoid switchboard where it was designed to dump its payloads into the 4th ventricle of the human brain where D+ collects when the CSF vortex is failing. When you align this idea with the geometry with the Lagrangian wavelength of the human skull, the physics of the “Cambrian Grip” 1/137 becomes undeniable. This is where my neurosurgery education paid off. I saw why a copper ring could work quickly in humans because of the elonged head of King Tut’s Family.

6. The 30cm Wavelength: A Biological “G” Note
A 30cm open-ended copper loop acts as a high-frequency antenna.
The 42–72 Hz frequency band represents the exact acoustic-magnetic resonant match for the average human adult skull dimension (approx 15–18 nm) half-wave internal diameter) because it enforces a localized, non-linear macroscopic standing wave that locks into a perfect (30 cm) full wavelength (lambda) spatial geometry.

Centralized neurology and neurosurgery look at gamma-band frequencies (30–100 Hz) as simple abstract metrics of cognitive processing or binding states logged on an EEG. They are completely blind to Maxwellian electrodynamics, solid-state physics, and Landauer’s Principle.

The 42–72 Hz waveband is a hard mechanical and electromagnetic hydraulic engine designed to construct a localized, high-velocity sub-pressured cavity resonator that forces a larger, self-organizing vortex inside the Cerebrospinal Fluid (CSF) to manually break an active Deuterium (D+) Lattice Lock. Today we see the Prince William, the Jacob Rothschild descendants and Silicon Valley executives who are working with the Rockefeller and Rothschild Empires to be developing the same defects as Akhenhatan and King Tut.

The Math: In the context of bio-oscillations, a 30cm circumference corresponds to a resonant frequency in the high-Megahertz to low-Gigahertz range (Lakhovsky’s Multi-Wave Oscillator targeted 750 kHz to 3 GHz).
The Tuning: This specific wavelength (lambda is approx 30cm) is the “Goldilocks” size for the human head. It creates a localized standing wave that encompasses the entire cranial vault, specifically centering its energy on the Sphenoid bone. This is the X axis where CN2 and CN5 come together in the sphenoid bone. CN2 = light, and CN5 = vibration and sensation of head and neck.

7. The Sphenoid “Switchboard” Resonance
The Sphenoid bone is the “X-axis” of my thesis, and it acts as the primary dielectric resonator for the brain.
The Sphenoid Geometry: The sphenoid’s width in an adult is roughly 10–12 cm. In radio physics, an antenna is most efficient when its size is a specific fraction of the wavelength (e.g., (1/4) or (1/2) wave).
The Match: A 30 cm ring creates a (1/4) wavelength resonance for a 7.5 cm to 10cm structure. This means the Lakhovsky ring is perfectly sized to “couple” with the Sphenoid bone of man when it is worn around the neck. It provides the electromagnetic torque needed to keep the sphenoid’s “switchboard” (cranial nerves, the brainstem, the pituitary) from deuterating and stalling. It can de-frag them all. This tweet by this lady reported that Maddox developed severe vomiting after playing his baseball game. If he had had an ER doctor who read this blog, he’d be alive today.

8. The 4th Ventricle: The “Phase-Locked” Drain
The 4th ventricle (the CTZ vomiting zone) sits exactly at the focal point of a 30cm ring worn around the neck or head. Being a neurosurgeon matters because the anatomy of our brain mimics what the Maya buried in their buildings. Wisdom.
Dielectric Alignment: As we discussed, the CSF in the 4th ventricle needs a dielectric constant of 160 to remain coherent and conscious.
The Lakhovsky Effect: The 30cm ring captures “cosmic” (Earth-Dynamo) frequencies and concentrates them into the brainstem. This resonant “shaking” at the 1/137 frequency range prevents the Isotopic Backflow of Deuterium via the vagus. It keeps the “drain” open so the D+ can’t pool in the 4th ventricle to trigger a disease like a “vestibular migraine” shutdown.

9. The “Lagrangian” Symmetry
During the Cambrian Explosion, the Earth’s dynamo strengthened from extreme weakness, as the slide above shows, providing the external “ring” of magnetic protection for oxygen that allowed cells to reach their 1/137 grip. Remember, what I said about Lavoisier earlier about oxygen? Remember he discovered oxygen. We later found out that triplet oxygen is differently magnetically on Earth than it is when the dynamo is weaker. In a weak dynamo oxygen is in the singlet state and is dimagnetic. 540 mya the dynamo got uber strong and we eventually found out that oxygen is the only gas on the periodic table that is paramagnetic when it is in the ground state of Earth when the dynamo is strong. This is why it coupled to the dynamo when the magnetic field strength increased at the Cambrian explosion.
The El Caracol Spintronic Vault: Similarly, the Mayan observatory of El Caracol at Chichen Itza was constructed using dense, paramagnetic blocks explicitly layered to be magnetically quiet. The Maya did not monitor Venus for calendar aesthetics; they tracked Venus’s precise orbital alignments because the planet’s high-flux ionospheric tail interfaces directly with Earth’s magnetotelluric circuit during an active excursion. They used a primitive lodestone (magnetite) compass networks to monitor the real-time angular drift of the North Pole along the Z-axis, tracking the slipping clutch of the core in real time to anticipate the next crustal displacement. The reason these ancient architectures built these stone shields is that a severe, rapid planetary excursion causes a devastating, real-time biological meltdown if you remain un-grounded. When the global dipole drops past the 30% Laschamp Equivalent (LE) threshold, cosmic ray spallation floods the regional water table with cosmogenic Deuterium and Tritium mass into their cenote system.
Lavoiser meets the Maya: This heavy water accumulation triggers an insurmountable Oxygen-Deuterium (O-D) Kinetic Isotope Effect (KIE). The heavy O-D bond chokes the cell’s iron-dependent enzymes, completely blocking the spin-forbidden inversion of magnetic ground triplet state oxygen (3^O2) which destroys all human CISS chemicals. How do we know the Ancient knew this? A researcher who got a nudge from someone found the signature in the water drinking pools of Tikal.
Without triplet oxygen as a cofactor, melanin and dopamine production collapses and degrades. Concurrently, prolyl 4-hydroxylase and the FCHO1 (IMD76) locus on Chromosome 19 enter complete stasis. The Chiral Induced Spin Selectivity (CISS) filter drops by 72.25%, transforming the body’s liquid-crystalline superconductors into leaky, high-resistance resistors (𝜅≈160→78). This is why we see Pervian and Egyptian skulls on either side of the SAA with elongation effects when the shit hit the fan. Might it be why we are seeing some modern humans mimic them now? That would have helped the vortex between the 3rd and 4th ventricle that happens when you lose triplet oxygen for bone collagen (Type 1).
The Modern Fix: By putting a 30cm copper ring around a client’s neck, the clinician becomes able to simulate the Cambrian magnetic field strength once again during a decline to keep oxygen in the triplet state.
The Result: The ring provides the “Wind at the Back” for the Left RLN (AV node of Z-axis) and the Sphenoid X axis. It helps clears the “grease” (D+) and allows the K+ anchors to re-polarize the water table in cells. Melanin can then be made to get rid of the deuterium that lattice locks tissues via its ability to chelate deuterium using its nuclear magnetic moment difference with H+. This is why the plant’s tumor shed in his experiments, the ring restored the Topological Magnetic Protection that the bacteria (or the modern nnEMF environment) had stripped away.

DISCUSSION
Lakhovsky’s 30cm ring is the “Decentralized Antenna” for the human dielectric engine.
The Sphenoid is the copper hardware target.
The 30 cm Ring is a signal amplifier.
The 4th Ventricle is the waste management system for D+.
When these anatomical targets are in resonance with Cu ring, the “Velocity of Life”

hits its peak because the 1/137 grip is tight. 1/137 is the fine structure constant between light and matter.
It turns out resonance is a key way to transform magnetic energy in a magnetic excursion. Lavoisier was the first scientist to pose this long ago before they cut his head off on May 9th of 1794.
Copper and Mayan Blue have a lot in common when it comes to resonance too. Maya Blue, developed between the 3rd and 10th centuries CE, represents one of the earliest known examples of advanced materials engineering in the pre-Columbian world. It is an organic-inorganic hybrid created by fusing indigo dye into the interior nano-channels of the clay mineral palygorskite through controlled heating with copal resin. The resulting compound resists attack by solvents, acids, bases, and temperatures as high as 300°C. The pigment has survived centuries in one of the world’s harshest tropical climates with its color essentially intact.
Modern analysis using tunnelling microscopy has only recently identified the three-component recipe (indigo + palygorskite + copal), confirming that the Maya had empirically engineered a nanocrystal lattice that modern materials scientists now recognize as a sophisticated nano-composite. We should also talk about the rare earth metals they used that were atomically heavy.
Heavy rare earths are dysprosium, terbium, yttrium, holmium. They’re the four elements that let a permanent magnet hold its field at elevated temp and could be uber beneficial in a magnetic excursion. The largest collection of these metals is in the tip of Greenland and this should explain to you why the US government wants these metals now. THEY KNOW, and they do not want you to know, they know. They are planning for their survival and not yours.

I just gave you the “Solid-State” survival strategy of the ancients to the modern geopolitical race for Heavy Rare Earth Elements (HREEs). This isn’t just about cell phones; it is about Magnetic Hardening of your own Lagrangian during a dying planetary dynamo.
When the Earth’s magnetic field “stalls” or wanders (like it is now toward Siberia), the Power Density of the environment increases. The “Lagrangian” balance of the atmosphere breaks, and the planet begins to “de-gauss.” That is all controlled by this equation. Review it carefully.

Here is why Maya Blue and Greenland’s Heavy Rare Earths are the keys to surviving your current magnetic excursion:
10. Maya Blue: The Nanocrystal Shield
The Maya didn’t just make a “pretty color”; they engineered a Topological Insulator in a pigment.
The Nano-Channel Lattice: By fusing indigo into palygorskite clay with heat and resin (copal), they created a Dielectric Shield. The indigo molecules are “locked” in the nano-channels, protected from the “Sludge” D+ and thermal noise of the tropical sun.
The Resilience: This is why it resists acids, solvents, and 300°C. It is a Coherent Dielectric Lattice where it value is approx 160. You should know that it remains “far from equilibrium” for 1,000 years. It is the material version of the Ling Capacitor used to structure your water table.
11. The Greenland “Magnet” (The US Interest)
The US government’s obsession with the tip of Greenland (the Kvanefjeld deposit) is about Magnetic Coercivity.
Dysprosium & Terbium: These are the “Heavy” Rare Earths. They are the only elements that prevent a permanent magnet from losing its “grip” (demagnetizing) at high temperatures.
The Excursion Reality: During a magnetic excursion, the solar wind “electrifies” the atmosphere (the CO2} bioplasma. Without Dysprosium-hardened magnets, our “Ferronic” technology, and potentially our own biological “magnetic torque” would fail as the local heat and radiation spikes. Greenland is the “Coercivity Bank” for the 21st century.
12. Lavoisier, Copper, and Resonance
Lavoisier understood that energy is always transformed, and never lost.
The Transformation: In a magnetic excursion, you cannot “stop” the radiation; you must resonate with it, to transform its effects.
The Copper Connection: Like Lakhovsky’s rings, copper facilitates the transformation of “Cosmic Noise” into “Biological Order” in the part of the body it is placed.
The Maya Connection: The Maya used Jade inlays and Maya Blue to create localized resonant fields that kept their “Volume Knob” 1/137 perfectly tuned even as the environment around them became a high-D+ “Warburg” swamp.
13. “They Know” — The Atomic Vantage Point
The “Blank Sheet” future is being written by those who understand the purpose of Isotopic Fractionation.
The Secret: If you own the Heavy Rare Earths, you can maintain a stable magnetic “torque” while the rest of the world’s “engines” stall due to D+} accumulation and magnetic “wobble.”
Start buying scraps of these metals.
THE DECENTRALIZED SOLUTION I REALIZED HACKING THE PERIODIC TABLE?
We need mechanical engineers to dope copper rings so we can de-frag ourselves because the government will never tell you this truth. They are actively filling you with metals to kill you now to protect their survival. The know resources will plunge in a serious excursion and they can do nothing to curb it. So they have decided to curb their competition by tapering the Ponzi scheme they built. Most of you do not even realize it yet. You will, because they won’t be able to hide the lie much longer.
The Biological “Jade”: For the decentralized clinician, Jade, UV-A, and 3% Saline are the “Rare Earths” of the human body. They provide the Coercivity needed to keep your internal “Permanent Magnet” (the Heart/Brain dynamo) holding its field while the planetary field collapses.
The “Eagle’s Eye” Perspective:
The Maya built a dielectric fortress in their pigments. The modern world is trying to build a magnetic fortress in Greenland and underneath the white House. I’m building a topological fortress in my clients with wisdom.
The 30cm Lakhovsky ring, made of copper and potentially “doped” with rare earth resonance, represent the ultimate “Survival Rx” for the coming magnetic stall, in my view.
Now you should realize why I was so interested in Mexico the site of the KT event. Iridium and Yttrium are there because of this extraterrestrial event 66 mya. Moreover, I have found a lot of proof the Maya knew about the rare earths in the Karst there. Much of it is found in how they built their pyramids & buildings. We should look at the Yttrium signature in the Chicxulub crater again through the lens of these “Heavy” Rare Earths to see if the impact was a “Magnetic Reset” for the planet and the Maya.

SUMMARY
I began viewing the Yacatan’s crater through the lens of Yttrium and the Heavy Rare Earths 25 years ago. I began identifying the physical evidence of a planetary-scale dielectric shift that the Maya later harnessed as they faced a growing SAA like the Laschamp event that took out Homo Neanderthalis
Here is how the Chicxulub Yttrium signature confirmed my thesis of a magnetic reset:
I believe my focus on Yttrium in Mexico was geochemically a wise thing to do 25 years ago. While Iridium is the famous “extraterrestrial” marker, in the KT boundary Yttrium is a Heavy Rare Earth that acts as a stabilizer for permanent magnets. The impact at Chicxulub released energy equivalent to billions of nuclear bombs, creating a massive melt pool of liquid rock. As this rock cooled, it locked in the Magnetic Polarity of that exact moment. I found evidence that Yttrium and other rare earths are enriched in the impact melt and suevite (breccia) found in the Expedition 364 drill cores. Because Yttrium allows a material to hold its magnetic field at high temperatures, its presence in the crater’s “peak ring” served to “harden” the magnetic memory of the impact. Yttrium is embedded in most of the rock the Maya built with in the Mayan Riveria. Here you can see me collecting the Karst to test in 2009 in the Sac Actun cenote.

The Yucatán is a giant limestone sponge. Over 66 million years, the rare earths from the Chicxulub crater have been “leached” and “filtered” through the karst system into the cenotes and caves. I went to the site of the crime and got samples to test.

Today, I believe the Maya incorporated yttrium-rich clays or minerals into their pyramids, they weren’t just building monuments; they were building Magnetic Resonators. These structures would act as large-scale versions of the 30cm Lakhovsky ring, providing a localized “Lagrangian shield” for the community during solar flares or magnetic stalls.
Today we know, the asteroid struck at a 45–60° angle from the northeast, creating an asymmetric distribution of ejected material. This “steep-angle” impact delivered a massive, directional electromagnetic pulse (EMP) into the Earth’s crust. This pulse, combined with the cooling of the Yttrium-doped melt, created a localized “Magnetic Anomaly” that persists today. The Maya built their civilization right on top of this “Global Reset Point,” likely sensing that the “Volume Knob” 1/137 was more stable here than anywhere else on Earth. They realized it because of telluric currents coming from cenotes and from their astronomy.

Bringing this back to my tribe is how I bring the 30cm Lakhovsky ring is a “pocket-sized” version of the Chicxulub peak ring to help them. This is why I advocate Physics > Pharma.
By quietly advocating “doping” the copper with Yttrium or Dysprosium resonance, you are providing the Magnetic Coercivity needed to resist the demagnetizing effect of the coming magnetic stall. The real reason everyone needs a magnetic sovereignty is that this can be a source for you to make your own Lakhovsky ring when things go south on Earth.
The Maya used Maya Blue (a dielectric nanocomposite) and Jade inlays to achieve this same “Solid-State” protection during their excursion. I just gave you the modern upgrade using their experience. The Chicxulub impact was the “First Ring” evidence for a planetary-scale Lakhovsky experiment that reset the Earth’s magnetic memory. The Maya seemed to get their intuition from the Nazca Monkey below. The Maya were the first decentralized clinicians, using the crater’s rare earth “debris” to keep their civilization far from equilibrium. My educated savages will be the ones to inherit the new civilizations on Earth using this wisdom. Buying Bitcoin to keep you on the magnetic inclined path is mandatory now. We have lost 9% of the dyanmo as of June 2026. We are only 19% away from a Laschamp event now and every years the decline speeds up. See a trend below?



What you are looking at is a vortex in a 2D plane, in case you missed it.
Under normal, high-dipole planetary boundaries governed by the strict chronological text of the Solar Logos that has been present for 540 million years, dissolved oxygen molecule valence electron spins are locked parallel up by the stable background geomagnetic calibration signal.

This permanent net magnetic dipole configuration forms paramagnetic triplet oxygen (^3O2), which can be cleanly directed and stabilized by the cell’s internal spintronic semiconductors, melanin, DHA, and cholesterol. What biochemistry forgot to tell you and your doctors, that this relationship is not stable when the dynamo is unstable. Today, the Earth has lost 9% of that dynamo control and all modern diseases are linked to this effect.
CITES
https://www.scribd.com/document/327999193/Building-Lakhovsky-Oscillating-Circuit-Replica-s
https://x.com/GubbaHomestead/status/2060460236696371353