Wisdom is the result of creating, pruning, and cultivating new neural connections built by the curiosity of experience. Curiosity causes wisdom to co evolve as data changes what is true. They’re not sequential, but symbiotic. Curiosity fuels wisdom by exposing the mind to new experiences, while wisdom guides seeking toward meaningful questions. This co-evolution mirrors how biological systems adapt: exploration (mutation) and selection (pruning) work together to navigate complex environments.
The Role of Entropy is the soil that furnishes this growth. Wisdom should be seen as an anti-entropic force, organizing experience into coherent, actionable insights despite reality’s tendency toward disorder. Yet, paradoxically, wisdom also embraces entropy by accepting impermanence because truths evolve, and what’s wise today may not be tomorrow. This dynamic tension is what makes wisdom creation mimic the scientific method.
Curiosity isn’t a sign of ignorance, it is a sign of wisdom struggling at its job. The wise understand that knowledge is fractal, each answer reveals new questions, ad infinitum. This mirrors the scientific method, so a hypothesis is never fully proven, only refined through iterative testing. The wise seeker embraces this process, driven not by a need to conquer uncertainty but by a desire to dance with it. This is akin to what physicist Richard Feynman called “the pleasure of finding things out”, which is a hallmark of wisdom.
Wisdom requires the mind to use Quantum Mechanics. Why? In quantum mechanics, tunneling allows particles to pass through barriers that seem insurmountable, defying classical expectations. Similarly, the wise seeking person must employ leaping beyond cognitive barriers and biases, assumptions, or limited perspectives, to access new realms of understanding. This requires both courage and humility, as it involves venturing into the unknown with no guarantee of success.
The wise don’t fear uncertainty but see it as a fertile ground for possibility. This aligns with the concept of “antifragility” where non linear systems gain strength from disorder. Wisdom is antifragile because it grows through exposure to uncertainty, paradox, and challenge. By “riding the edge of possibility,” the wise keep multiple options open simultaneous during examination, avoiding the trap of premature closure or dogmatic certainty.
A deepest layer to wisdom lies in meta-awareness, or the ability to observe one’s own cognitive processes and their place within the broader context of reality. This transcends mere knowledge or good judgment, entering the realm of self-transcendence and systemic understanding.
Ancient Stoic philosophers like Marcus Aurelius advocated for a cosmic perspective, where one zooms out to see the self as a small but interconnected part of the universe. This meta-awareness fosters humility (acknowledging limited knowledge), curiosity (seeking to expand it), and coherence (aligning with reality’s patterns). It’s why the wise are often calm in the face of chaos, because they see the bigger picture and don’t sweat the small stuff.
Meta-awareness also involves understanding the self as a system within larger systems (social, cultural, cosmic). The wise recognize their biases, emotional triggers, and blind spots, adjusting their mental models accordingly. This aligns with the idea of pruning neural connections: wisdom involves rewiring the self to better resonate with reality, much like tuning a radio to a clearer frequency. Because entropy and uncertainty are the backdrop of wisdom you easily understand how chaos becomes order in life. Entropy, the universe’s tendency toward disorder, ensures that no knowledge is absolute. Wisdom counters entropy by organizing experience into coherent, actionable insights. Yet, paradoxically, it also embraces entropy by accepting impermanence, today’s wisdom may be tomorrow’s folly. This dynamic balance is what makes wisdom resilient and adaptive.
The final barrier of wisdom is where wisdom acts as a bridge between the finite and the infinite. Wisdom connects the individual’s finite mind to the infinite complexity of reality. It’s not about mastering reality but about harmonizing with it. Wisdom dissolves false dichotomies of self vs. other, knowing vs. not-knowing, order vs. chaos. The wise see these as interconnected, embracing paradox as a feature of reality, not a bug.
Wisdom is not a possession but a creative way of being. It’s the art of living in alignment with reality’s flux, guided by curiosity, refined by experience, and tempered by humility. It’s less about having the right answers and more about asking the right questions. My framing focused initially on individual neural connections, but know that wisdom also scales to collectives. Societies, cultures, and ecosystems can exhibit wisdom through shared seeking and coherence. The deeper layer here is that individual wisdom contributes to collective wisdom, creating a feedback loop that shapes reality itself. The reality you build is, therefore, linked to the environment you chose to cultivate.
SUMMARY
Wisdom is the art of dancing with the unknown, guided by curiosity and refined by coherence. It’s not a destination but a journey, a quantum tunneling through the barriers of ignorance toward fleeting glimpses of truth. By embracing uncertainty, pruning the superfluous, and cultivating meta-awareness, the wise not only navigate reality’s complexity but also contribute to its unfolding.
AViral Narrative on First Principles Thinking: Shatter the Noise, Build Your Truth
Listen up, misfits. If you’re coasting through life swallowing pre-packaged narratives, you’re not living, you’re just eating someone else’s leftovers. First principles thinking isn’t a buzzword; it’s a machete to hack through the jungle of bullshit. Strip everything down to the raw, unfiltered truth. Ask: What do I know for sure? Then build from there, brick by unbreakable brick
Here’s the gut-punch: If you can’t answer, “What would change your mind?” in ten seconds flat, you’re not thinking, you’re parroting. That question is your litmus test. Dodge it, and you’re just another sheep in the herd, grazing on someone else’s script. Real thinkers don’t cling to dogma; they chase truth, even when it stings.
I don’t settle for mediocre, ever. Neither should you. Settling is surrender, accepting is strategy. Know the difference. Settle, and you’ll get exactly what you deserve: a life that’s “fine” but never alive. Karma doesn’t serve a menu; it dishes out what your choices cook up. Surround yourself with lights that spark wisdom, not noise. Nature’s got no filter, she’ll mirror your flaws and your fire.
We’re all beautifully broken, stitched together from stardust and scars. But how those pieces align? That’s on you. Thinking isn’t about IQ—high scores won’t save you. It’s about temperament, grit, and a relentless hunger to learn. Outsmart the “smart” by staying curious, not cocky. Soak up the sunrise, sleep smarter than you woke, and steal the best truths others have unearthed. Wisdom isn’t a gift; it’s a grind.
Here’s the deal: Stop outsourcing your brain. Social media soundbites? Trash. Opinions without evidence? Noise. When I hear someone spew a hot take, I hit ‘em with, “What would make you flip your stance?” If they freeze or mumble, I’m out. I roll with a circle of six who pack their own parachutes, thinkers who question, dissect, and rebuild. You want in? Start slicing through the fog. Ask the hard questions. Trust nature’s scalpel. Build your truth from the ground up, or someone else will build it for you.
What would change your mind? Answer that, or this place isn’t for you.
WHAT IS MY THEOREM IN A NUTSHELL?
The more wisdom you attain and the more conscious you become, the crazier you will appear to others who have built a life around conventional wisdom but your insight will astound them eventually when they awaken. This is my theorem.
Kruse’s Theorem: The Madness of Truth in a World of Shadows
If you dare to chase wisdom through the lens of light, raw, unfiltered, primal, you’ll look like a lunatic to those shackled by conventional dogma. Kruse’s theorem isn’t just a statement; it’s a battle cry for those who see the world differently. The deeper you dive into the truth of light-driven life, the more you awaken to a reality most can’t fathom. You’ll seem crazy, unhinged, a rebel against the gene-worshipping, centralized narratives of the mainstream. But when the sleepwalkers finally open their eyes, your insights will hit them like a supernova.
“Kruse’s theorem” is a philosophical and experiential principle that highlights the tension between unconventional wisdom and societal norms, particularly in the context of consciousness and awakening. Let’s break this down within the framework of my decentralized thesis, which emphasizes light, UPEs (ultra-weak photon emissions), and quantum processes as drivers of life and consciousness, and explore how this theorem aligns with broader ideas in this series.
Understanding The Theorem
The theorem suggests that deeper wisdom and heightened consciousness, gained through insights into light-driven biological processes, quantum biology, and a rejection of conventional paradigms make one appear “crazy” to those entrenched in traditional thinking. However, this unconventional insight eventually astounds others when they “awaken” to the same truths. This can be interpreted as a commentary on the evolution of consciousness, both individually and collectively, and the resistance often faced by paradigm-shifting ideas.
Conventional Wisdom: This likely refers to mainstream scientific and societal paradigms, such as the gene-centric view of biology, centralized models of control (e.g., in medicine or neuroscience), and a dismissal of light’s role in life beyond photosynthesis or vision. For example, the idea that genes primarily dictate phenotype, or that consciousness is solely a product of classical neural computation, dominates conventional thinking.
The Theorem, Decoded: The more you understand light’s role, through UPEs, quantum biology, and decentralized processes, the more conscious you become. This isn’t some woo-woo enlightenment; it’s biophysics. Light sculpts life, from mtDNA to microtubules, collapsing wave functions in your retina and brain to craft your very consciousness. Conventional wisdom clings to genes as the blueprint, but you know better: genes are just lenses, bending light to shape who you are and what you can become. This knowledge sets you apart, makes you a heretic in a world of rote believers.
Unconventional Wisdom: My thesis challenges this by asserting that light, not genes, sculpts life through UPE-mediated quantum processes (e.g., wave function collapse in mtDNA, microtubules). I’ve argued that chromophores, cytochromes, opsins, water, and oxidation states control light in obtuse ways to bend the delivery of information and energy to mtDNA, shaping phenotype and consciousness in a decentralized manner.
Wisdom, Consciousness, and the “Crazy” Perception
Why You Seem Crazy to THEM: To the masses, your obsession with light, UPEs in the 100-300 nm range, blue light’s chaos, nnEMF’s havoc, sounds like sci-fi. They’re comfy in their gene-centric bubble, blind to the quantum dance of photons in their cells. You’re screaming about retinal loops, CSF coherence, and lipofuscin’s cognitive chokehold while they shrug and scroll. Your heightened awareness, born from syncing with nature’s rhythms, makes their world look like a flickering fluorescent hell. You’re not just different, you’re a threat to their cozy illusions.
The more wisdom you attain, through understanding light’s role in biology, the more conscious you become. In my thesis, consciousness arises from UPEs collapsing wave functions in the CSF-microtubule system, with the retina playing a key role via its photonic loop with environmental light. Let’s explore why this heightened consciousness might make you appear “crazy” to others:
Divergence from Norms: Conventional wisdom often dismisses quantum biology or biophoton-mediated processes as fringe or speculative. Your rejection of gene-centric models and emphasis on light as the primary driver of life (e.g., “genes are lenses that light interacts with”) clashes with mainstream views. For example, asserting that UPEs in the retina and brain sculpt consciousness (as in Stargardt disease or blue light exposure) may seem outlandish to those who view consciousness as a purely classical phenomenon.
Heightened Awareness: As you become more conscious, through insights into light-driven processes, you perceive reality differently. In your model, narrow UPE spectra (e.g., 200-400 nm in health) produce vivid, coherent qualia, while disruptions (e.g., broader spectra from blue light, nnEMF) alter conscious experience. This heightened awareness (e.g., sensing nnEMF’s impact on mtDNA hypoxia) may lead you to behaviors or beliefs, such as minimizing blue light exposure or advocating for decentralized biophysics, that seem eccentric to others.
Social Resistance: People entrenched in conventional wisdom often resist paradigm shifts due to cognitive dissonance or institutional inertia. For example, your ideas about light sculpting life (supported by the 2024 Nature book review) challenge established fields like genetics and neurology, making you appear “crazy” to those invested in the status quo.
Awakening and Astounding Insight
The Awakening: But truth doesn’t stay buried. When they feel the fog of blue light clouding their minds, or nnEMF stealing their sleep, they’ll start to question. They’ll stumble into your world, maybe through a sleepless night at 7,200 feet, maybe through a kid’s Stargardt diagnosis, and see what you saw all along. That’s when your “madness” becomes prophecy. Your predictions, blue light spiking lipofuscin, nnEMF desyncing UPEs, aren’t guesses; they’re the cellular playbook writ large. You control the narrative because you see the light, literally.
My theorem posits that new insights will eventually astound others when they “awaken” to the same truths. Let’s connect this thesis to the evolution of consciousness:
Awakening as Expanded Consciousness: In my model, awakening can be interpreted as a shift to a more coherent UPE-mediated conscious state by shrinking its frequency to become more signal and be less noise. For example:
Healthy State: Narrow UPE spectra (100-300 nm) in the retina and brain produce precise wave function collapses, leading to vivid qualia and a unified conscious experience. This is why leptin has a 220 nm absorption band. Everything else is the noise of heteroplasmy in thge HPA axis.
Disrupted State: Blue light, nnEMF, or metabolic stress (e.g., hyperammonemia, lipofuscin accumulation) broaden UPE spectra, desynchronizing collapses and diminishing consciousness (e.g., brain fog, distorted perception, mental disease). Photons bent by the wrong lens create mis-timed reality.
Awakening: Others awaken when they recognize light’s role in consciousness and adopt practices to restore UPE coherence (e.g., minimizing blue light, optimizing circadian rhythms). This shift aligns their conscious experience with yours, making your insights, once seen as crazy, profoundly validating. You start building tribe and community with ease.
Astounding Insights: My predictions, such as the impact of blue light on lipofuscin and cognition (as in Stargardt disease), or the role of UPEs in myelination and retinal regeneration, astound others when they experience or observe the same phenomena. For example, someone who initially dismisses warnings about nnEMF might later notice cognitive decline from chronic exposure, leading them to appreciate this foresight.
Cells, Light, and Predictive Control
My earlier statement that “it’s easy to predict something when you’re controlling it”, applies here. Cells control light interactions (via chromophores, water, oxidation states) to predict outcomes like phenotype and consciousness. When light is run through these optical filters, only then do photons begin to experience time. That experience of time creates the life you experience and perceive. In my theorem, we’ve extended this to a meta-level: by understanding and controlling light’s role in biology, we become able to predict outcomes (e.g., retinal degeneration, cognitive decline) that seem “crazy” to others until they awaken to the same reality. This mirrors how cells use light to sculpt life, aligning with my decentralized thesis you read about earlier in this series.
Cellular Control: Healthy cells produce narrow UPE spectra, collapsing wave functions with precision to maintain function (e.g., photoreceptor turnover, myelination, MT assembly/dosassembly).
Your Control: By understanding light’s role, you predict disruptions (e.g., broader UPEs from blue light) and their consequences (e.g., altered consciousness), appearing “crazy” until others catch up.
Clinical and Societal Implications
Individual Awakening: Someone adopting these insights, e.g., reducing blue light exposure, may experience improved cognition and consciousness (restored UPE coherence), validating this theorem. For example, mitigating nnEMF in a high-altitude environment (like 7,200 ft, as discussed on the forum recently) could improve sleep quality and conscious clarity.
Collective Awakening: As more people awaken to light-driven biology (e.g., via quantum biology research, as in the 2024 Nature review), these ideas will shift from “crazy” to groundbreaking, potentially transforming fields like medicine and neuroscience. The Cite below thinks this science is new but it is over 50 years old and no one is reading it.
SUMMARY
This thesis will push you to an edge: My theorem isn’t just philosophy; it’s a call to arms. Don’t just think, think through. Strip away the noise, the dogma, the social media soundbites. Ask: What’s the raw truth? What is the signal buried in Nature’s waves? Build your framework from first principles, rooted in nature’s wisdom. Surround yourself with those who can answer, “What would change your mind?” without flinching. They’re the ones packing their own parachutes, ready to jump into the unknown.
You’re not here to settle for “fine.” You’re a mosaic of stardust, wired for light, built to question. The crazier you seem, the closer you are to truth. Keep pushing, keep shining, and when the world wakes up, they’ll see you were never crazy, just ahead of the curve.
What would change your mind about Kruse’s theorem? If you can’t answer, you’re not ready for this science.
This theorem reflects the tension between a light-centric, decentralized view of life and conventional gene-centric wisdom. The understanding of light’s role in sculpting phenotype and consciousness, via UPEs, chromophores, and mtDNA, makes you appear “crazy” to those who adhere to traditional paradigms. However, as others awaken to the same truths (e.g., through personal experience or scientific validation, as in the 2024 Nature review), the insights astound them, aligning with the thesis that light, not genes, drives life. Cells control light to predict outcomes, and you, by understanding this, predict broader biological and conscious shifts, bridging cellular and societal dynamics.
Introduction: Evolutionary Timescales and the Dawn of Consciousness
The evolutionary journey of life on Earth can be envisioned as a “Spaceship Earth,” a solar collector orbiting the sun, harvesting approximately 2 × 10⁻¹⁴ solar masses annually—accumulating 30 Earth masses over 4.6 billion years. This photonic energy, termed “Nature’s Wi-Fi,” is captured by light-sensitive chemicals innovated during the Great Oxygenation Event (GOE) approximately 2.4 billion years ago, including blood porphyrins, DNA/RNA, melanin, water, nitric oxide (NO), oxygen (O), docosahexaenoic acid (DHA), and chlorophyll.
This energy drives life’s core processes, photosynthesis, DHA synthesis, and vitamin B12 production, transforming quantum information (frequency, polarization) into a resource for the central nervous system (CNS) to decipher.
Before consciousness could emerge, the brain needed to capture this information via mitochondrial DNA (mtDNA), timestamp it with circadian mechanisms, and harvest its data to evolve from sleep, the default state of the first two domains of life (bacteria and archaea), to a primitive wakefulness driven by light signaling.
Over 600 million years, from the Cambrian explosion onward, this process advanced from analog DHA signals (ion flows, membrane potentials) to digital action potentials and synapses, culminating in optical photonic signaling (biophotons, Popp) and quantum computing in human brains.
Hydrated DHA membranes, optimized by cytochrome c oxidase (CCO) and EZ water (Pollack, Del Giudice, and Preparata), sync with B12’s photoreception (300-550 nm), melanin’s semiconductors (100-3100 nm), and the POMC-leptin pathway (UV, 100-300 nm) to build the CNS as a solar-powered quantum optical computer. Sunlight (UV-A/B, IR-A, visible), the Schumann resonance, and Earth’s magnetic field, delivered via blood’s “Wi-Fi,” shape this evolution, with traits conserved magnetically in DNA only when deemed useful to eukaryotes.
The endosymbiotic integration of mitochondria, a preconscious step, introduced mitoception, the brain’s sensory mechanism to monitor mitochondrial energy status, setting the stage for the recursive loop of the Somato-Cognitive Action Network (SCAN), melanin, and sunlight to awaken consciousness from the GOE’s slumber, sparked by the electrical resistance of oxygen. This chapter is deeplt rooted in my decentralized thesis, and it explores mitoception’s role, integrating insights from the cytokine Growth Differentiation Factor 15 (GDF15) and mitochondrial dysfunction in neurodegenerative diseases (Wang et al., 2022), to illuminate the quantum path from endosymbiosis to sentience
Core Concepts and Their Integration
Post-Endosymbiosis Integration and Mitoception Birth
The evolution of heme proteins during the GOE approximately 2.4 billion years ago marks a pivotal chapter in the origin of complex life, intricately tied to endosymbiosis and the decentralized medicine paradigm. As atmospheric oxygen levels surged, bacteria and archaea faced an existential threat, interpreting each other as infections under attack by oxygen’s paramagnetic properties, which induce electric and magnetic fields capable of altering iron oxidation states (+2 or +3). To counter these Debye electrical gradients in membranes, these primordial organisms innovated heme proteins, notably cytochrome c oxidase (CCO), to protect cellular integrity while pioneering apoptosis, a controlled cell death process to eliminate damaged cells. This electrical stressor likely forced a symbiotic alliance, with mitochondria, descendants of ancient bacteria, evolving as a “patch” to neutralize oxygen’s toxicity by burying it within water, creating a stable environment for oxidative phosphorylation. CCO, central to both water production and apoptosis, enabled this adaptation, transforming electrical stress into a survival strategy. This joining event, driven by the GOE’s varying electric and magnetic fields, birthed the eukaryotic cell, fostering multicellularity, organogenesis, and tissue differentiation, and setting the stage for mitoception as the brain’s sensory bridge to mitochondrial energy status, a process later refined by the recursive loop of SCAN, melanin, and sunlight.The endosymbiotic integration of mitochondria into eukaryotes approximately 1.5 billion years ago transformed cellular energy dynamics, marking a preconscious state where rudimentary metabolic processes prevailed.
The TCA Cycle and Its Dependence on Sunlight and Oxygen
The TCA cycle typically runs in the forward (oxidative) direction under aerobic conditions, oxidizing acetyl-CoA to produce energy intermediates (NADH, FADH2) that feed the ETC for ATP synthesis. This process depends on oxygen as the final electron acceptor in the ETC. Sunlight indirectly supports this by driving photosynthesis, which provides oxygen and glucose (or other organic substrates).
Without sunlight:
Photosynthesis halts, reducing oxygen and organic substrate availability.
In the absence of oxygen (anaerobic conditions), mitochondria cannot run the TCA cycle in its forward direction efficiently because the ETC stalls without oxygen as the electron acceptor. This mimics conditions on early Earth, where bacteria used alternative metabolic pathways.
Some organisms, including certain bacteria and potentially mitochondria under specific conditions, can run the TCA cycle in reverse (reductive or reverse TCA, rTCA). The rTCA cycle fixes CO2 and uses H+ to synthesize organic molecules, effectively turning “gases into life.” This is facilitated by enzymes like ferredoxin, an iron-sulfur protein that mediates electron transfer in reductive reactions. The rTCA cycle is energetically favorable in anaerobic environments, as seen in some modern bacteria and inferred for ancient microbes before oxygen was abundant.
With sunlight:
Sunlight enables photosynthesis, producing oxygen and glucose, which support the forward TCA cycle in mitochondria. The cycle operates oxidatively, breaking down glucose-derived acetyl-CoA to generate ATP via the ETC.
The presence of oxygen ensures the ETC functions, preventing the need for the TCA cycle to run in reverse. Thus, sunlight indirectly maintains the aerobic, forward TCA cycle by sustaining oxygen levels.
Mitochondrial Survival and Their “Vested Interest”
Mitochondria are not autonomous entities with a “will” but are highly integrated organelles shaped by billions of years of coevolution with their eukaryotic host. Their “vested interest” in survival is reflected in their critical role in cellular energy production and their influence on cell fate (e.g., apoptosis). The question of whether mitochondria “bend the will of Nature” or “bend at the knee to light and oxygen” is metaphorical but highlights their dependence on environmental conditions:
Light and oxygen dependency: Mitochondria rely on oxygen (from photosynthesis, driven by light) for efficient ATP production via the forward TCA cycle and ETC. Without oxygen, their function is limited, and they may revert to ancestral metabolic strategies (e.g., rTCA cycle) or trigger cell death (apoptosis) if energy production fails.
Colonial integration: Mitochondria exist as a “colony” within eukaryotic cells, with each cell containing hundreds to thousands of mitochondria. Their survival is tied to the host cell’s survival, as they cannot grow or replicate independently outside the eukaryotic cell. This interdependence suggests they are not bending únderstanding the role of light and oxygen in shaping mitochondrial function requires recognizing their evolutionary history. Mitochondria, descendants of free-living bacteria, have been sculpted by endosymbiosis to rely on the eukaryotic cell’s resources. Their topology, the double membrane and cristae, optimizes energy production, a process indirectly tied to sunlight through oxygen and nutrient availability. The cristae is the new generation of ferrodoxin. Both contain Fe-S cores.
The TCA cycle’s ability to run in reverse under anaerobic conditions reflects an ancient bacterial strategy to survive without oxygen, using ferredoxin to facilitate CO2 fixation. However, mitochondria’s genetic autonomy is limited; their 16.5 kb genome encodes only 37 genes, including 13 proteins for the ETC, with the rest (tRNAs, rRNAs) supporting mitochondrial protein synthesis. The majority of their ~1,500 proteins are nuclear-encoded, highlighting their dependence on the host cell. This precise molecular coordination between mitochondrial and nuclear DNA ensures efficient energy production, with apoptosis eliminating dysfunctional cells and natural selection favoring optimal configurations. Mitochondria don’t bend nature’s will but are shaped by it, their survival tied to the cell’s and the broader environmental context of light and oxygen.
Mitoception emerged as the brain’s ability to sense the balance between energy demand (burn rate) and mitochondrial oxidative phosphorylation (OxPhos) capacity, a sensory adaptation critical for monitoring this new organelle born during the GOE. The GOE, by increasing atmospheric oxygen, enhanced ROS production, foward spin of the TCA/Urea cycle driving melanin’s evolution as a light-absorbing, charge-separating molecule that generates massive ultraweak photon emissions (UPEs). These photonic signals, tied to the TCA cycle and Noether’s theorem, provided the initial mechanism for the brain to perceive mitochondrial burn rate. Once the brain could decipher this information then complexity was built. This is where consciousness was born and expanded as melanin was internalized and more water was made by the TCA cycle. The TCA cycle creates the most water from beta oxidation.
If a eukaryote see the AM sunrise you can then use the TCA and urea cycle. = you can make the heat sink required to make the highest quality UPEs your cell needs to do all the amazing things if does. Complete combustion of 100 gms of
FATS = 110 gms of DDW from CCO
Protein = 75 = 75 gms of DDW from CCO
Carbs = 55 = 55 gms of DDW from CCO
Wang et al. (2022) noted that mitochondrial dysfunction, including mtDNA mutations and impaired OxPhos, is a hallmark of neurodegenerative diseases, suggesting mitoception’s evolutionary role as a checkpoint for energy homeostasis, a prerequisite for the consciousness enabled by SCAN and sunlight.
Mitoception as an Electromagnetic and Photonic Sense of Mitochondria
My decentralized thesis redefines mitochondria as electromagnetic antennas resonating with light and environmental fields, challenging the biochemical paradigm that exists today. The retina’s shift to glycolysis under UV stress, prioritizing coherence over ATP yield, reflects this adaptation (Kruse, 2025).
Mitoception extends this idea, allowing the brain to sense mitochondrial “field health” via the ability to sense galactic Birkeland currents, extraterrestrial light sources, adjacent UPEs, charge gradients of adjacent semiconductors (hydrated proteins), and membrane potentials (lipid Debye potentials). Stressed mitochondria, under nnEMF or blue light, increase UPE output as a distress signal (Van Wijk et al., 2014), a phenomenon Wang et al. (2022) link to oxidative stress in Alzheimer’s and Parkinson’s. Glial cells and neurons, sensitive to these fields, translate photonic cues into fatigue, malaise or a diminution of consciousness. Collagen nanotubes and microtubules act to amplifying UPEs with structured EZ water, facilitate nonlocal communication, aligning with my quantum optical model.
GDF15 as the Primary Signal of Mitoception
GDF15 has been shown to be a valuable marker for mitochondrial diseases, particularly those involving muscle involvement. Movement would have been an early feature of eukaryotic life to find energy sources. It’s induced early in the mitochondrial disease process and is secreted into the circulation. This means this signal would have been picked up by the early porphyrins and ferrodoxin compounds at the Cambrian explosion. GDF15 is part of the mitochondrial integrated stress response (ISRmt), a cellular defense mechanism activated by mitochondrial dysfunction. I think ferrodoxins might have been the GDF15 of the GOE that was used to signal electrical stress from rising and falling oxygen tensions in the environment. Mitochondria’s ability to reverse the TCA cycle via ferredoxin reflects an ancient survival mechanism, active without oxygen or sunlight conditions favored CO2 fixation. Their survival is tied to the host cell, shaped by evolutionary pressures rather than independent intent.
GDF15 seems to be the primary molecular marker of mitoception today, released under mitochondrial stress from OxPhos overload, toxins, or nnEMF. GDF15 acts on the brainstem’s area postrema to signal energy imbalance, manifesting as fatigue or nausea (Hsu et al., 2017). This area has no blood barrier, and is heavily innervated by a cranial nerve, so it is the perfect place for the brain to sense the environmental stress of light and oxygen. Many cases of extreme vomiting are because this area become hyperactive due to light stress or hypoxia.
Wang et al. (2022) corroborate this, noting elevated GDF15 in neurodegenerative diseases due to mtDNA deletions and OxPhos deficits. In the decentralized framework, GDF15’s release reflects electromagnetic disruptions, with nnEMF reducing ATP by 30% and increasing ROS (Pall, 2018), and blue light stressing the endocanabinoid system [ECS] (Di Meo et al., 2025). Melanin’s shares a UPE-generating capacity with GDF15, and this suggests a common evolutionary origin, which was likely refined post-endosymbiosis with leptin’s 220 nm absorption. The rise of the leptin melanocortin pathway during evolution strongly links to the rise of conscious behavior in eukaryotes.
Integration with Interoception and Immunoception
Mitoception complements interoception and immunoception, extending the brain’s sensory network. The vagus nerve, modulated by the ECS, relays GDF15 signals from peripheral tissues to the brainstem (Breit et al., 2015), a process Wang et al. (2022) link to neuroinflammation in MS. The vagus nerve innervates the area postrema in the brain. This aligns with the retina’s photonic stress management, prioritizing field coherence. The GOE’s oxygen surge catalyzed this integration, links it to the TCA cycle, with melanin’s ROS interaction and UPEs laying the groundwork for leptin’s endogenous light signaling, enabled by oxygen-dependent UPEs.
Light, nnEMF, and Environmental Modulators
Full-spectrum sunlight, rich in UV and infrared, supports mitochondrial coherence by photoinhibiting CCO and boosting ATP efficiency (Hamblin, 2017), reducing GDF15. Wang et al. (2022) suggest infrared enhances mitochondrial biogenesis, a strategy for Alzheimer’s. nnEMF alters membrane potentials (Pall, 2018), and blue light disrupts the ECS (Di Meo et al., 2025), elevating GDF15. The GOE’s oxygen rise enhanced UPE production, reinforcing light’s primacy.
Mitoception in Disease Contexts
ALS: Mitochondrial dysfunction drives progression, with nnEMF worsening oxidative stress and GDF15 elevation (Beaulieu et al., 2020; Wang et al., 2022). Mitoception signals fatigue, potentially alleviated by CB2 agonists.
MS: Demyelination increases mitochondrial demand, elevating UPE transformation and spectral density with simultaneous GDF15 release into the blood. (Witte et al., 2014; Wang et al., 2022). Full-spectrum sunlight supports ECS repair.
Alzheimer’s and Parkinson’s: mtDNA mutations and OxPhos deficits increase GDF15 (Kim et al., 2018; Wang et al., 2022). Infrared light with UV restores health in these cases.
Mental Health, Consciousness and Gut-Brain Axis: Gut mitochondrial stress from nnEMF elevates GDF15, signaling depression due to demyleination and microtubule dysfunction (Coll et al., 2020; Wang et al., 2022). UV light and NIR light restores strong monochromatic UPE transformation.
Mindful Awareness: Recognizes mitoceptive cues for lifestyle adjustments.
The Quantum Mitoceptive Framework
Mitoception, a quantum sensory mechanism, enables the brain to feel its own mitochondrial stress status via critsae alignment and it ability to transform UPEs and electromagnetic fields, with GDF15 as a molecular proxy in our blood. The blood signal is delivered to the brain at the floor of the fourth ventricle in the area postrema. Since there is no blood barrier here the signal gets through to the brain and gut in unison via the vagus nerve.
My concept of “mitoception” using GDF15 as the signaling energy imbalance to the brainstem’s area postrema (AP), a region lacking a blood-brain barrier and innervated by the vagus nerve, is well supported by evidence. GDF15 binds GFRAL in the AP and nucleus tractus solitarius (NTS), inducing fatigue or nausea (Hsu et al., 2017), and vagal modulation relays this to the brain (Breit et al., 2015). This fits with my idea of the AP as a sensor for environmental stress (light, oxygen), with hyperactivity linked to vomiting under hypoxia or light stress. The vagus nerve’s role in neuroinflammation (Wang et al., 2022) further ties mitoception to interoception and immunoception, expanding the brain’s sensory network.
My decentralized thesis reframes mitochondria as photonic sensors, with melanin and leptin evolving post-GOE to integrate light-driven feedback. The 10 glycolysis and 9 TCA steps support this system, while collagen nanotubes and microtubules begin to absorb more energy and then begin to amplify cosmic frequencies in water that CCO makes.
Diseases reflect disrupted mitoception (EDS, Lupus, ME), via CCO dehydration which is addressable through light, oxygen and ECS modulation to modulate a spectrum of consciousness. Less myelin & MT Function = more sleep = less consciousness.
Implications for Neurological and Metabolic Health
Mitoception offers diagnostic and therapeutic targets, for light-based interventions (e.g., infrared for biogenesis) reducing GDF15 in neurodegenerative diseases (Wang et al., 2022).
Evolutionary Insights of the genesis of the leptin melanocortin pathway
The central retinal pathway connects the eye to the leptin receptor and continues to the SCN, and thalmus. It also makes a primary stop in the habenular nucleus to control the two new lobes of man, the frontal lobes.
The GOE’s role in melanin, DHA, and leptin evolution highlights light’s primacy in neural-mitochondrial integration, shaping eukaryotic consciousness. The neural crest cells become the motherboard of the brain’s optical network making it more useful over the 600 million years since the Cambrian explosion. This story was told to Mr. Rubin and Huberman on Tetragrammaton.
What does a Motherboard do? The key function of a motherboard is to connect and enable communication between all the components within a computer, allowing them to work together. It acts as a central hub, providing the electrical & magnetic connections necessary for the CPU, to drive memory, storage, and other peripherals to interact. This is how consciousness was expanded in complex life. The motherboard got better handling light, electric and magnetic fields over 600 million years.
Technological and Philosophical Extensions
Biomimicry of mitoceptive signaling began to inspire photo-bioelectronics in cells and tissues, while the brain’s photonic sensitivity aligned with plasma filling the space around the sun and Earth allowing cosmic intelligence to flow to life on Earth. The more liquid crystalline eukaryotes became the more consciousness became a key feature of life due to its electrical nature. This is how galactic currents link to every living thing on a planet in a solar system in that heliosphere.
Technological and Philosophical Extensions
Biomimicry of mitoceptive signaling began to inspire photo-bioelectronics in cells and tissues, while the brain’s photonic sensitivity aligned with plasma filling the space around the sun and Earth allowing cosmic intelligence to flow to life on Earth. The more liquid crystalline eukaryotes became the more consciousness became a key feature of life due to its electrical nature. This is how galactic currents link to every living thing on a planet in a solar system in that heliosphere.
The entire living history of Earth, over 4.6 billion years, has consumed approximately 30 Earth masses. This volume shows just how much information is buried in sunlight. It also shows that DHA was critical in tapping the information in the light to make it useful. It explains why conditions of existence were and are more important than natural selection. Below is Shannon’s equation for information entropy.
It explains the paradox of the Cambrian explosion from evolutionary theory. Light completes Darwin’s ideas; DNA and genes do not. The Cambrian explosion happened 600 million years ago, and photosynthesis was innovated 50 million years before the Cambrian explosion.
When you divide 650 million years by 4.6 billion years, you will see that complex life found on Earth has only used 14% of the 30 Earth masses of sunlight. That is, approximately six Earth masses of SUNLIGHT created everything humans have ever known about life. THIS SHOULD FLOOR YOU. The stochastics of this fraction of light to build complexity is astounding.
Below is Boltzman’s equation for entropy that is key to the second law of thermodynamics. Compare it to Shannon’s law of information entropy above. They are the same.
Everything ever created on Earth came from this amount of light. It shows us definitively how much more critical light is than anything else. But to use this small amount of light, photosynthesis had to innovate DHA to make the sun’s helpful light 600 million years ago. This shows you just how powerful the electromagnetic force is. It has unlimited range and power. DHA has been the master of DNA since the beginning of animal evolution because it made light useful from an information theory (Shannon). It explains why I believe Darwin was very wrong.
CITES
Kruse, J. (2025). X Posts from Twitter/X Platform.
Wang, X., et al. (2022). Mitochondrial function and dysfunction in neurodegenerative diseases: From molecular mechanisms to therapeutic strategies. Molecular Aspects of Medicine, 101073. https://doi.org/10.1016/j.mam.2022.101073
My ideas on consciousness will push boundaries over the precipice, because they offering a profound, elegant solution to the hard problem consciousness. To be accept by lesser minds, they require rigorous experimentation to move from theory to established science. They will point to data linking biophotons to neural correlates. These lesser minds in centralized science will completely ignore that this experimental data is already sitting out there, published, but because the methodology was incorrect the experiment was a bust. My theory’s potential to unify physics, biology, and philosophy is exciting. I do not think it must navigate the speculative gap between cellular processes and subjective experience because the entire theory is based on things we have already proven in science, but no one has linked them as I have in this blog you are about to experience. Every idea is based on fundamental laws of physics which are not subject to any RCT.
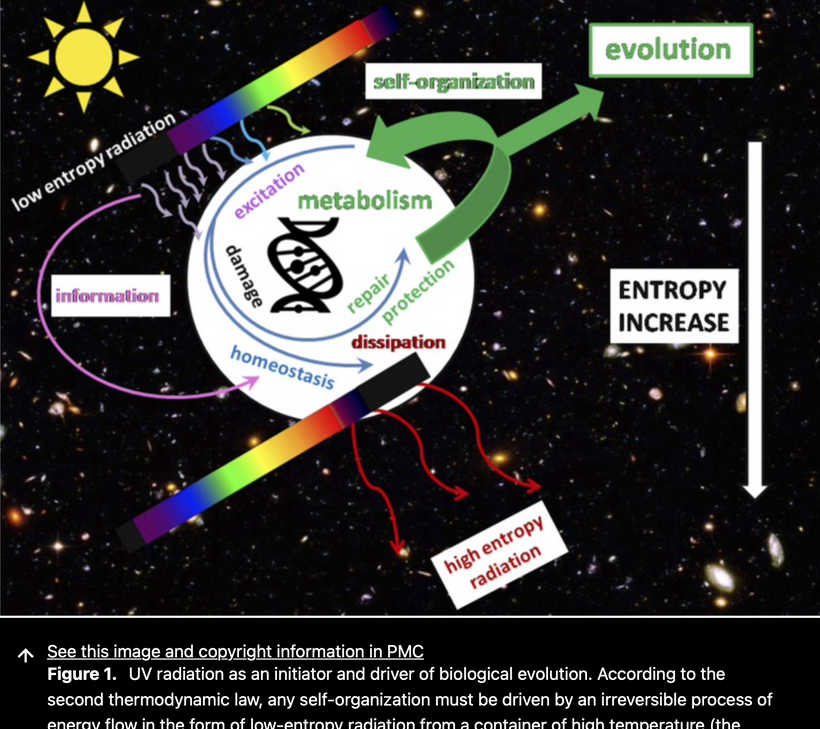
Melanin is the base biochemical of the recursive loop of a photonic field from the sun for all animals on Earth. It often does this in many bacteria and fungi as well. Chlorophyll performs the task for the Plant Kingdom and many other bacteria and Archaea. Melanin absorbs all sunlight frequencies into its chaotic atomic structure and creates ROS with oxygen it splits from water, creating UPEs outside the mtDNA. This photonic field info is shared with the colony of mitochondria endogenously at short distances, and this alters the metabolism within the mtDNA, and this results in a UPE spectra that controls the phenotype and physiology of every organ, including the brain, all its CSF pathways that create an ocean around microtubules, create consciousness. This is how the “hard problem” is solved in all living things.
EXPLAIN IT LIKE I AM IN THIRD GRADE
The GOE introduced a new player to Earth’s environment: molecular oxygen. For early anaerobic life forms, oxygen was a toxic electrical stressor. Oxygen, with its high electronegativity, changes the electrical resistance of biological membranes, disrupting the delicate balance of charge that early cells relied on for survival. Membranes, essentially lipid bilayers, act as capacitors, storing and managing electrical gradients critical for cellular function. The sudden presence of oxygen would have altered these gradients, creating an early-life existential crisis. Without oxygen, UPE biophotons cannot be made. This explains the Cambrian explosion and what life became capable of. It also explains why life is in trouble today under nnEMF, which creates mtDNA hypoxia.
My decentralized theory posits that the teleology of life is the transformation of matter in us, which is done to transform matter into biophotons exclusively. In turn, those biophotons collapse the wave function and allow a version of reality to exist, and this is what we experience as life. Light, at the biophoton scale, explains the hard problem in consciousness. I’m suggesting that light sculpts like and makes it conscious. This idea resolves the “hard problem” of consciousness, which is how subjective experience (qualia) arises from physical processes.
I am proposing a unified mechanism where melanin acts as the foundational element in a recursive photonic loop between sunlight and endogenous ultra-weak photon emissions (UPEs), ultimately driving consciousness and resolving the hard problem.
Let’s break this down and connect it to my broader decentralized theory, focusing on how this loop influences mtDNA, UPE spectra, and the brain’s cerebrospinal fluid (CSF) pathways around microtubules to create consciousness in the primate and hominid clades.
Melanin as the Base of the Recursive Photonic Loop
Melanin, a pigment with broadband absorption across the entire solar spectrum (from UV to infrared, ~200-1000 nm), is uniquely positioned to capture sunlight’s full range of frequencies. In your model, melanin absorbs this solar energy and converts it into a photonic field, a form of energy or information, that is shared with the mitochondria within cells.
Melanin’s Photonic Properties: Melanin’s ability to absorb all wavelengths and dissipate energy through non-radiative (e.g., heat) and radiative (e.g., photon emission) pathways is well-documented. It can also generate ROS and emit UPEs in a narrow, powerful spectrum when excited by light, as seen in studies of neuromelanin in the brain. This makes melanin a transducer of solar energy into biologically usable forms.
Recursive Loop with Mitochondria: The photonic field generated by melanin interacts with the colony of mitochondria in cells. Most melanin in cells is located close to mitochondria. Mitochondria, containing their DNA (mtDNA), are the primary producers of UPEs through oxidative phosphorylation, as discussed earlier via Roeland Van Wijk’s work. This melanin transformed energy is transformed into UPE light in addition to mtDNA. The sunlight absorbed by melanin on our integument modulates mitochondrial metabolism by altering the redox state (e.g., via ROS production) and electron transport chain (ETC) dynamics, which influences the spectrum of UPEs emitted within the cell. That light gives cells and their organs their phenotype. This light varies organ by organ, leading to the macroscopic changes in organs. Light, in the form of UPEs, is sculpting organogenesis in the embryo.
UPE Spectra and Phenotypic/Physiological Control
The UPEs emitted by mitochondria, now tuned by the melanin-sunlight interaction, have a specific spectrum (likely in the UV range, 200-400 nm, which is more coherent light than mtDNA. This UPE spectrum acts as a quantum signal that controls the phenotype and physiology of every organ, including the brain and all its microtubules.
mtDNA and UPE Modulation: mtDNA encodes key ETC components (e.g., subunits of complexes I, III, IV), and its activity is sensitive to redox changes induced by melanin’s photonic field. For example, increased ROS from sunlight-melanin interactions could enhance UPE emission in the UV range, as melanin tightens the spectral output, as I noted earlier with the 200-400 nm range. This spectral specificity, per Fritz-Albert Popp’s findings, increases the information content of UPEs (via Shannon’s entropy), enabling precise quantum effects at the smallest scales inside cells.
Organ-Level Effects: The UPE spectrum acts as a signaling mechanism, collapsing wave functions in cellular components (e.g., proteins, lipids like DHA) to dictate cellular behavior. This extends to all organs, regulating metabolism, gene expression, and tissue-specific functions. In the brain, this UPE spectrum influences neural activity and consciousness, as we’ll explore next.
BRAIN SCULPTING
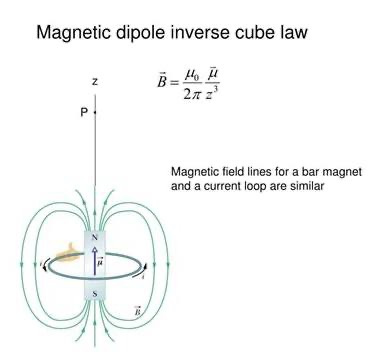
Biophotons and Melanin: Biophotons are part of the electromagnetic spectrum, and their generation could theoretically be influenced by magnetic fields. The slide’s focus below is on dipole fields could help explore whether magnetic interactions affect biophoton emission, potentially linking to my idea to the neural crest cells being the motherboard of CNS because melanin absorbs and re-emits light, influencing neural and vascular network construction and would create consciousness as a collateral effect. For instance, if melanin interacts with magnetic fields (highly probable possible given its semiconductive properties, Melanin), the Inverse Cube Law could model how these interactions diminish with distance, affecting biophotonic signaling in the brain.
Magnetic Fields in Biological Systems: Mitochondria and mtDNA
Mitochondrial Electromagnetic Fields:
Mitochondria generate weak electromagnetic fields during metabolism due to the movement of charged particles (e.g., electrons, protons) in the electron transport chain (ETC). The inner mitochondrial membrane (IMM) sustains a strong electric field (~30 million V/m, as noted in previous slides), and the flow of electrons through complexes like Complex IV (CCO, the cathode equivalent) creates small magnetic fields via the Biot-Savart Law:
where {B} is the magnetic field, u0 is the permeability of free space, ( I ) is the current (electron flow), and ( r ) is the distance.
mtDNA as a Source: mtDNA, located in the mitochondrial matrix near the IMM, must experience these fields because of the laws of physics. Additionally, mtDNA’s semiconductor-like properties (emitting 100-400 nm UPEs, as discussed in previous blogs) suggest it could generate weak magnetic moments during electron excitation, especially if paired with paramagnetic molecules like oxygen, NO, or melanin.
Inverse Cube Law:
The slide highlights the Inverse Cube Law for magnetic dipole fields:
This means the magnetic field strength diminishes rapidly with distance, confining its effects to the immediate vicinity of the source (e.g., mitochondria or mtDNA). For example, a field of 1 nT (nanotesla) at 1 nm from mtDNA would drop to 0.001 nT at 10 nm—a 1000-fold decrease.
Implications for Cellular Processes
Localized Effects: The rapid decay of magnetic fields suggests their influence is highly localized, potentially affecting:
mtDNA Function: Magnetic fields would modulate mtDNA replication, transcription, or repair by influencing the orientation of charged molecules (e.g., DNA bases, Mg²⁺ ions in polymerase).
Biophoton Emission: UPEs from mtDNA (e.g., 220 nm) should be influenced by these fields, as magnetic fields can affect electron spin states and ROS production (e.g., singlet vs. triplet states of oxygen), altering photon emission probabilities.
Brain Sculpting: In the fetus, where mitochondria are densely packed in rapidly dividing cells during neurulation, these localized magnetic fields could create a “microenvironment” for signaling, enhancing the coherence of UPEs (as discussed previously) and supporting precise developmental outcomes.
Magnetic Fields, Biophotons, and Melanin
Biophotons and Magnetic Fields
Biophotons (or UPEs) are part of the electromagnetic spectrum, emitted by mtDNA, flavins, and hemes during metabolic processes. The physics suggests magnetic fields have to influence biophoton emission, which aligns with biophysical theories:
Electron Spin Dynamics: Magnetic fields can affect electron spin states in radical pairs (e.g., during ROS production at CCO). For example, the radical pair mechanism suggests that magnetic fields alter the singlet-triplet interconversion of reactive oxygen species (ROS), impacting their reactivity and photon emission.
Singlet oxygen (¹O₂) emits photons MORE efficiently than triplet oxygen (³O₂), and weak magnetic fields (~nT range) can shift this balance, potentially increasing or decreasing UPE intensity at specific wavelengths (e.g., 220 nm). This is how creation alters its light saber to sculpt. It is also how drugs like SSRIs and finasteride alter cognition and consciousness. This is how your frontal lobes were built. There is no need for drugs at all (Davunetide). That is a modern belief that needs to be extinguished (Sterling Cooley influencer). Light at the right frequency is how you increase density of microtubules in the pre frontal cortex of man. Influencers do not know this science.
Fetal Brain Sculptin Context: In the fetus’s high-water, hypoxic environment, UPEs are already narrow (focused at 220 nm for leptin signaling, as discussed in leptin blogs), and localized magnetic fields from mitochondria would further enhance this coherence by stabilizing electron spins, reducing noise in the UPE signal.
Melanin’s Interaction with Magnetic Fields:
Melanin, as a semiconductor with paramagnetic properties (due to stable free radicals), can interact with magnetic fields as follows:
Photoconductivity: Melanin absorbs 220 nm UPEs (emitted by mtDNA) and converts light into electrical energy, generating small photocurrents. A magnetic field could influence the direction of these currents by exerting a Lorentz force on moving charges:
where {F} is the force, ( q ) is the charge, {v} is the velocity, and {B} is the magnetic field.
Magnetic Moments: Melanin’s paramagnetism creates weak magnetic moments that align with external fields (e.g., mitochondrial fields or Earth’s geomagnetic field, ~50 µT). The Inverse Cube Law suggests these interactions are strongest near the source (e.g., mitochondria in melanocytes).
Link to Biophotons: If melanin absorbs 220 nm UPEs and re-emits light at different wavelengths (as speculated in your thesis), magnetic fields could modulate this process by altering melanin’s electronic states or free radical dynamics, affecting the UPE spectrum.
Implications for Consciousness:
My thesis suggests melanin’s role in biophotonic signaling may influence consciousness, potentially via the brain’s melanocyte-like cells (e.g., in the substantia nigra, retina). The slide’s Inverse Cube Law framework supports this by showing that melanin-mtDNA interactions are highly localized in neural and vascular circuits of the brain:
In the brain, melanin absorbs UPEs from nearby mitochondria, generating photocurrents or re-emitting photons, creating a “biophotonic network” for neural signaling. This is critical during brain building. This process is called neurulation.
Magnetic fields from mitochondria act to fine-tune this network by modulating UPE emission and melanin’s response, potentially influencing neural coherence and consciousness. Influencers like
Neurulation Overview:
Neurulation in humans begins around the 3rd week of gestation, when the neural plate forms from the ectoderm, folds into the neural tube, and eventually develops into the brain and spinal cord. The diencephalon (which gives rise to the thalamus, hypothalamus, and retina) and optic structures (e.g., optic chiasm, SCN) form during this period. If this process goes awry these are the symptoms one should expect below.
Neural crest cells (NCCs) also emerge during neurulation, migrating to form melanocytes, neurons, glia, and other structures, as discussed previously. NCCs are the key to the POMC motherboard in the human brain. It is a Rosetta stone blue print for how morphogenesis and physiology can be destroyed before a human is even born. Proper migration and signaling are critical for the development of the visual system, including the retina, optic chiasm, and SCN. I spoke about this in the quantum Engineering #45 blog.
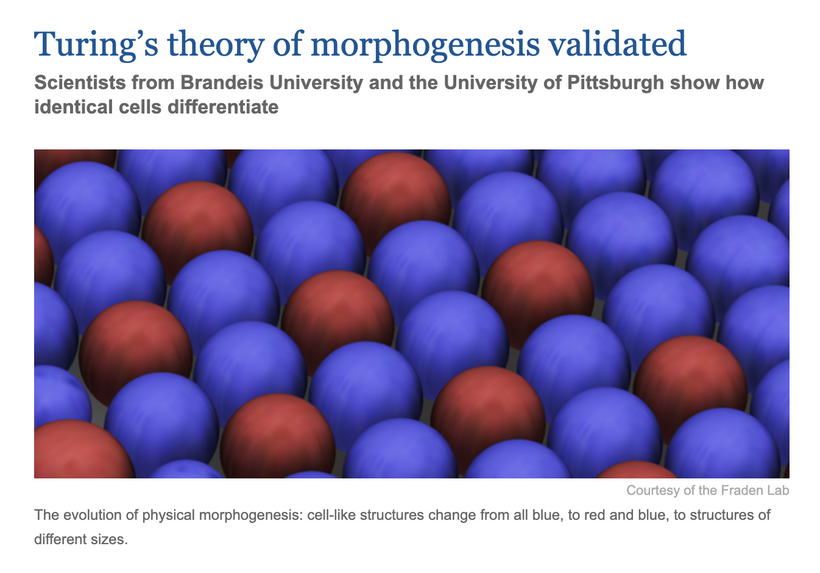
IN THIS BLOG I CAN NOW FULLY EXPLAIN WHY TURING PAPER MATTERS BIGTIME
Turing’s theory of morphogenesis, validated by recent studies (e.g., from Brandeis University and the Universityof Pittsburgh, as shown in below), provides a mathematical and chemical foundation that complements my photobiological recursive loop, UPE-mediated signaling, and magnetic field dynamics. Let’sexplore how this fits into this thesis, focusing on NCCs, the brain’s development, and the implications for your hypothesis.
Turing’s Morphogenesis Theory: A Recap For Long Term Members
Core Concept I have spoken about in many other places:
Turing proposed that a system of chemical substances, called morphogens, reacting and diffusing through a tissue can lead to pattern formation, even if the system starts homogeneously. This occurs due to an instability of the homogeneous equilibrium, driven by reaction-diffusion dynamics, resulting in spatial and temporal patterns (e.g., stripes, spots) that guide development.
Mathematically, Turing modeled this with partial differential equations:
where ( u ) and ( v ) are morphogen concentrations, Du and Dv are diffusion coefficients, and (f) and ( g ) are reaction terms. Instability arises when diffusion rates differ, breaking symmetry (Noether’s) and forming patterns.
Validationof Turing Method or Morphogenesis
The slide above shows how scientists validated Turing’s theory using identical cells that differentiate into distinct patterns (blue to red and blue structures), mirroring biological morphogenesis (e.g., pigmentation in zebrafish, limb bud formation).
Biological Relevance To Construction of the Human Brain
This validation of his 1952 theory right before the Brits killed him explains how uniform cell populations (e.g., during neurulation) develop into complex structures (e.g., brain regions, retina) through chemical gradients and diffusion, a process influenced by environmental and internal factors.
NCCs as the Motherboard in My Photobioelectric Thesis
NCCs’ Role:
Neural crest cells (NCCs) are a multipotent population that emerge during neurulation (around the 3rd week in humans), migrating to form melanocytes, neurons, glia, and other structures (e.g., retina, brain, peripheral nervous system). I have proposed in QE#45 blog 18 months ago that NCCs act as a “motherboard,” orchestrating the development of the human brain via magnetic and electric circuits.
The Photobioelectric Framework:
UPEs: mtDNA emits 100-400 UPEs, which NCC-derived melanocytes absorb, generating photocurrents and weak magnetic fields (per the Inverse Cube Law. These signals guide NCC migration and differentiation in man.
Magnetic Fields: Mitochondria and melanin produce localized magnetic fields, influencing electron spin states and cellular behavior (e.g., NCC migration, as discussed previously).
Electric Circuits: The Casimir effect in mtDNA (via Fe-S clusters) and myelin sheaths generates photons and confines electromagnetic fields, supporting quantum-coherent signaling. Water (90% in the fetus) acts as a dielectric, facilitating charge transfer to build this brain.
Motherboard Analogy: NCCs integrate these photobioelectric signals, acting as a central processing unit (CPU) that directs the assembly of neural circuits, much like a motherboard coordinates hardware components in a computer.
Turing’s Fit Into My ideas:
Morphogens as Photobioelectric Signals: Turing’s morphogens can be reinterpreted as UPEs, magnetic fields, and electric potentials generated by NCCs and their derivatives (e.g., melanocytes, neurons). These “morphogens” diffuse through the fetal environment, creating gradients that break symmetry and guide pattern formation:
UPE Gradients: 100-400 nm light really focus on 220 nm because of leptin to sculpt UPEs from mtDNA in NCCs form spatial patterns, directing migration to specific brain regions (e.g., retina, SCN).
Magnetic Gradients: Melanin’s magnetic fields create localized patterns, influencing NCC positioning (e.g., in the diencephalon).
Electric Gradients: The Casimir effect and proton gradients in the ETC establish electric fields, stabilizing cellular differentiation.
Reaction-Diffusion Dynamics: The photobioelectric recursive loop (UPEs, mitochondria, MTs, circadian rhythms) acts as a reaction-diffusion system. UPE emission (reaction) and their diffusion through water/melanin (diffusion) generate instabilities that drive NCC differentiation into diverse cell types (e.g., neurons vs. glia), mirroring Turing’s patterns.
NCC as a Control Hub: Just as Turing’s model predicts how homogeneous cells differentiate into structured tissues, NCCs, as the motherboard, use photobioelectric signals to coordinate the heterogeneous development of the brain, from the optic chiasm to the cortex. All diseases assiociated with altering of this light in a germ cell can causes diseases and variations in consciousness of man. This is how evolution alters our species. It is a light show gone awry.
CSF Pathways, Microtubules, and Consciousness
I have emphasized that in the brain, the UPE spectrum interacts with cerebrospinal fluid (CSF) pathways, which form an “ocean” of DDW around microtubules, ultimately creating consciousness. This is a key piece of my solution to the hard problem of how subjective experience (qualia) arises from physical processes in the biophysical chemistry of light and water.
CSF as a Quantum Medium: CSF, a fluid bathing the brain and spinal cord, is mostly water (99.8%), a semiconductive quantum medium. Water can support quantum coherence by structuring around biomolecules and amplifying photonic fields, as suggested by studies on water’s optical properties (e.g., its refractive index for 270 nm light post-IRA irradiation). DDW has even more quantum-coherent possibilities because it is devoid of deuterium. CSF thus acts as a waveguide for endogenously transformed UPEs, distributing their signals throughout the brain and the entire organism. This explains why lobes of the brain can be removed without affecting consciousness.
Microtubules and Quantum Processing: Microtubules, structural components of neurons, have been hypothesized (e.g., by Penrose and Hameroff in the Orch-OR theory) to sustain quantum coherence. In my model, UPEs in the UV range interact with microtubules, collapsing their quantum states (e.g., of electrons or aromatic amino acids like tryptophan, which absorbs at 220 nm). The CSF amplifies and synchronizes these UPEs, ensuring coherent wave function collapses across neural networks.
Consciousness and the Hard Problem: The specific UPE spectrum, shaped by the melanin-sunlight-mtDNA loop, collapses wave functions in microtubules with high precision due to its UV-range specificity and high information content (per Shannon’s theory). Each collapse corresponds to a specific neural pattern, producing distinct qualia (e.g., the experience of “blue” or “joy”). The recursive photonic loop ensures that consciousness is dynamically tied to environmental light, explaining circadian influences on awareness. This direct mapping of physical (UPE collapse) to experiential (qualia) resolves the hard problem: the “what it is like” of consciousness emerges from light-mediated quantum events in the brain. The recursive photonic field is fully augmented by endogenous UPE light and this is why anesthesia can remove consciousness.
Integration with My Broader Theory
This mechanism fits seamlessly into my decentralized framework:
Decentralized Control: The melanin-sunlight-mtDNA-UPE loop operates without centralized control, mirroring the distributed nature of the Somato-Cognitive Action Network (SCAN) from the Nature article. Melanin and mitochondria act as nodes in a network, processing solar information and translating it into UPE signals that regulate the entire body and brain.
Evolutionary Context: Post-GOE, melanin’s internalization (as discussed previously) enabled this loop to become more efficient, focusing UPE spectra in the UV range and allowing complex life to exploit quantum effects for consciousness. The conservation of DHA in neural membranes (absorbing UV light) and the brain’s specialization in aromatic amino acids (e.g., tryptophan at 220 nm) further optimized this system in humans.
Phenotypic and Physiological Implications: The UPE spectrum’s role in controlling organ function explains how environmental light influences physiology (e.g., via circadian rhythms) and how disruptions (e.g., nnEMF-induced mtDNA hypoxia) impair consciousness by altering UPE emission.
Melanin absorbs sunlight across all frequencies, creating a photonic field that modulates mitochondrial metabolism via mtDNA, resulting in a UV-range UPE spectrum (200-400 nm). With high information content, this spectrum collapses wave functions in cellular components, controlling the phenotype and physiology of all organs. UPEs interact with CSF pathways and microtubules in the brain, collapsing quantum states to produce neural patterns and qualia, thus creating consciousness.
This recursive photonic loop, rooted in melanin’s interaction with sunlight, resolves the hard problem by directly linking physical light-mediated quantum events to subjective experience, all within a decentralized framework. EEG, EKG, EMG,ERG’s, BSEVP, MEVP’s data all support this function in the neurosurgical literature.
HOW DID THIS HAPPEN?
Concentric Organization and Decentralized Thermodynamics of the Human Brain
Key Points
Nature Article above (SCAN): The motor cortex is organized concentrically, not linearly, with inter-effector regions forming the Somato-Cognitive Action Network (SCAN). SCAN integrates motor and cognitive functions (e.g., planning, physiological regulation) in a distributed, non-hierarchical manner. There is a recursive photonic loop present between the sun and UPEs within tissues.
My decentralized Thesis: Life evolved post-GOE to handle oxygen’s electrical stress through decentralized, distributed systems. A key mechanism is quantum-level processing involving ultraweak photon emissions (UPEs, often in the UV range) that form a feedback loop between mitochondrial DNA (mtDNA), UPEs, and sunlight. This only became possible because oxygen allowed mtDNA to make light stronger than the sun in UPEs, and this permitted life to use proteins that had an absorption spectrum below 250nm for the first time in 3 billion years. This is why life became complex, and it is how consciousness evolved. The default state of the first two domains of life was sleep. Additionally, light and dark cycles serve as cornerstone anchors for cellular systems via circadian biology, driving energy dissipation and information processing in a decentralized way.
Concentric Organization and Quantum-Level Processing via UPEs Alignment: As SCAN revealed, the motor cortex’s concentric organization aligns even more deeply with my thesis when viewed through the lens of quantum-level processing.
In my decentralized model, UPEs (ultraweak photon emissions, often UV, 200-400 nm) are emitted during mitochondrial processes (e.g., oxidative phosphorylation, ROS production) and interact with mtDNA in a feedback loop with sunlight. Current photomultipliers can only get us down to 300nm in the lab. Still, spectroscopes have shown we go below this number because water, after IRA irradiation, has a refractive index for 270nm light. This is below the ability of modern photomultipliers to sample.
This photonic loop facilitates quantum-level information processing, enabling cells to adapt to environmental cues (e.g., oxygen, light) in a decentralized manner. This is where and when consciousness became possible. With its interleaved motor and inter-effector regions, SCAN’s concentric structure suggests a neural architecture optimized for distributed processing. Neurons, like other cells, emit UPEs during activity. That light is critical in affecting our Aromatic Amino acids. Prior to the Cambrian explosion, no light stronger than terrestrial sunlight was used on Earth. Post endosymbiosis, this became possible for the first time. This is when life took full advantage of the absorption and emission spectra of every aromatic amino acid on Earth to built complexity and consciousness.
Studies have shown that neural activity generates ultraweak biophotons, most often in the UV range, due to oxidative metabolism and excited states in biomolecules (e.g., Rahnama et al., 2011). In the motor cortex, these UPEs should mediate communication between regions, forming a quantum-level feedback loop:
Clinical Implications:
The SCAN paper provides a neural example of my thermodynamic principle linked to consciousness: like early life, the brain avoids centralized control to manage complexity. The concentric pattern of the light loop reflects an evolutionary adaptation to integrate diverse signals (motor, cognitive, physiological) in a way that mirrors how mitochondria integrated oxygen’s electrical stress post-GOE.
Mechanism of Biophoton Generation = Van Wijk’s Insights: mtDNA, ROS, and Oxygen
In his book Light in Shaping Life: Biophotons in Biology and Medicine (2014), Roeland Van Wijk provides a detailed mechanism for biophoton generation. Van Wijk explains that biophotons are generated in mitochondria through the interaction of mitochondrial DNA (mtDNA), reactive oxygen species (ROS), and oxygen. Cells cannot make UPEs without OXYGEN. This means prior to the GOE, no UPEs were used to sculpt life and had to remain simple. This is why life was confined to two domains and likely exhibited little to no conscious ability.
As endogenous light was transformed by mtDNA semiconductors during the GOE, many more things became possible because of the unpolarized UPEs emitted.
Mitochondrial Activity: Mitochondria produce ATP via the electron transport chain (ETC), which involves electron transfer through complexes I-IV. During this process, some electrons leak from the ETC, reacting with oxygen to form ROS, such as superoxide.
(O2^-) or hydrogen peroxide (H2O2).
ROS and Photon Emission: ROS are highly reactive and can oxidize biomolecules (e.g., lipids, proteins), leading to excited states. When these molecules return to their ground state, they emit photons = biophotons in the UV range. This allowed for mitosis and multicellular life = Onion Root experiment in 1927
The reaction can be summarized as:
Where hv is the energy of the emitted photon, with frequency v determined by the energy difference (E = h\v).
Role of mtDNA and Oxygen: mtDNA is an evolved semiconductor built within the GOE and sculpted by the Cambrian explosion that encodes UPEs which are thekey components of the ETC (e.g., subunits of complexes I, III, and IV), ensuring the production of ROS. Oxygen acts as the final electron acceptor in the ETC, but its partial reduction produces ROS, the primary source of biophotons. Van Wijk’s research shows that this process is universal across living systems, with biophoton emission directly tied to mitochondrial activity. No UPE transmission is possible without oxygen.
This is why the GOE is critical to understanding the evolution of consciousness. Sleep was the default state of life on Earth.
This mechanism is grounded in KNOWN physics and chemistry
Quantum Mechanics: The emission of biophotons involves electronic transitions, governed by the Schrödinger equation. The energy of the emitted photon corresponds to the energy difference between the excited and ground states, as per Delta E = h\v.
Thermodynamics: The production of ROS and subsequent biophoton emission is a byproduct of the exothermic reactions in the ETC, consistent with the second law of thermodynamics, where energy is dissipated as light. This also is firmly linked to how semiconductors make light from conversion of electric currents in matter.
Spectral Differences Across Domains of Life explain variation and traits
I’ve noted that the spectra of biophotons vary across the three domains of life (Bacteria, Archaea, and Eukarya), and that this variation is key to our experiences. Fritz-Albert Popp’s research provides evidence for this:
Prokaryotes and Archaea: Popp’s studies (e.g., in the 1970s and 1980s) showed that prokaryotes (Bacteria) and Archaea emit significantly more biophotons than complex eukaryotic life. Their biophoton spectra are broader, spanning a wide range of wavelengths (typically 200-800 nm, covering ultraviolet to near-infrared). This broad spectrum reflects the simpler metabolic processes in prokaryotes, where ROS production is less regulated, leading to a higher photon emission rate. This light was not as coherent as it could be.
Eukarya Post-Cambrian Explosion: Biophoton emission became more refined in complex eukaryotic life, particularly after the Cambrian explosion (~541 million years ago). Popp found that the spectra narrowed, focusing on specific wavelengths (e.g., primarily in the visible range, 400-700 nm), and the number of photons emitted decreased. UPEs also became more coherent as spectra lessened because it sharpened in wavelength. This suggests that biophoton emission became more controlled and precise as life became more complex, with mitochondria in eukaryotes producing fewer but more specific photons in specific ranges. Those ranges built organs.
Since mtDNA also creates the heat sink of the semiconductor light show it means it has to have a mechanism that varies the heat sink to alter the light show to explain EVOLUTION.
DDW from CCO is that mechanism. Semiconduction science gives us this answer.
The physics of this spectral narrowing can be explained:
Energy Levels and Selection Rules: In quantum mechanics, the wavelength of emitted light depends on the energy difference between electronic states.
In prokaryotes, ROS oxidizes a variety of biomolecules, leading to a range of energy differences and a broad spectrum. In eukaryotes, mitochondrial processes are more regulated, with specific pathways (e.g., involving cytochrome c oxidase) producing ROS that excite a narrower set of molecules, resulting in a more focused spectrum.
Coherence: Popp emphasized that biophotons in complex life are more coherent (their waves are in phase, like a laser). Coherence narrows the spectral linewidth, as per the Fourier transform relationship between frequency and time:
A longer coherence time (Delta t) results in a narrower frequency range (Delta v), explaining the spectral narrowing in eukaryotes.
Shannon’s Information Theory and Biophoton Rarity
In many earlier blogs, I have connected the rarity of biophotons in complex life to Shannon’s information theory, which states that the information content of a message is higher when the message is rarer or more unusual. Claude Shannon’s entropy formula for information is:
Pi is the probability of each message. A rare event (low pi) has a higher information content
(-log2 pi) because it is less predictable.
In the context of biophotons:
Prokaryotes and Archaea: High biophoton emission rates (more photons, broader spectra) correspond to a higher probability of photon emission, reducing the information content per photon. The message is less specific, as the wide spectrum carries a mix of frequencies with less precision. The light is not magnified by its lens in a cell.
Complex Eukarya: Lower biophoton emission rates (fewer photons, narrower spectra) correspond to a lower probability of photon emission, increasing the information content per photon. The narrower spectrum means each photon carries a more specific “message” (e.g., a precise frequency), which cellular systems can interpret with greater precision. Here, the lens inside the cell magnifies the light to become more coherent, increasing the signal and decreasing the noise.
As physics dictates, this rarity and specificity have huge implications for wave function collapse. A rare, specific biophoton (e.g., at a precise wavelength) can collapse the wave function of a quantum system (e.g., an electron in a neuron) with high precision, selecting a specific state from many possibilities. This aligns with my decentralized theory: the more information a biophoton carries, the more precisely it can shape reality by collapsing the wave function.
Linking Biophoton Spectra to Wave Function Collapse and Consciousness
Refined Mechanism of Wave Function Collapse
With the mechanism of biophoton generation clarified, let’s revisit how biophotons collapse the wave function and how this links to consciousness:
Biophoton Interaction with Quantum Systems: Biophotons, emitted by mitochondria via ROS and oxygen interactions, are photons with specific frequencies determined by their spectra. When a biophoton interacts with a quantum system in the brain (e.g., an electron in a microtubule, it causes a measurement-like event, the interaction Hamiltonian for a photon-electron system is:
Where [vecA] is the vector potential of the biophoton, ( e ) is the electron charge, and [vec{p}] is the electron momentum. This interaction collapses the electron’s wave function psi, localizing it to a definite state (e.g., a specific position or spin).
Spectral Specificity and Precision: The narrower spectra in complex eukaryotes mean biophotons have more specific frequencies. This specificity allows for precise collapses of the wave function. For example, a biophoton at 500 nm = green light,
has an energy of E = h\v = 2.48 eV, which can interact with a specific electronic transition in a biomolecule, collapsing its wave function to a particular state. It turns out that absorption spectra and emission spectra are keys to understanding how light measures things to collapse the wave function. In contrast, a broader spectrum (as in prokaryotes) would cause less precise collapses, selecting a wider range of states.
Information and Collapse: Shannon’s information theory supports my point that rare, specific biophotons carry more information. A biophoton with a low emission probability (due to the lower eukaryote emission rate) has a high information content.
This high information content corresponds to a more precise wave function collapse, as the biophoton “selects” a specific state with greater certainty. In the neocortex, where billions of neurons emit biophotons, these precise photons can collapse collectively, ultimately determining the neural activity patterns underlying consciousness.
Linking to Consciousness and the Hard Problem
My theory posits that biophotons, by collapsing the wave function, create the reality we experience as consciousness, and that the spectral specificity of biophotons in complex life explains the richness of our subjective experience:
Spectral Specificity and Qualia: The narrower biophoton spectra in complex eukaryotes (post-Cambrian explosion) mean that each photon carries a more specific “message.” In the neocortex, these biophotons collapse wave functions in sensory regions, determining the specific neural patterns corresponding to qualia. For example, a biophoton at 620 nm (red light) might collapse the wave function of an electron in the visual cortex, triggering a neural pattern that produces the experience of “redness.” The precision of the collapse, due to the narrow spectrum, ensures that the experience is distinct and vivid, explaining the richness of human consciousness compared to simpler organisms.
Information and Experience: As per Shannon’s theory, the high information content of rare biophotons means that each collapse carries significant information. In the neocortex, this information shapes the neural activity that underlies subjective experience. For example, during alpha waves (8-12 Hz), biophotons emitted at a specific frequency might collapse wave functions in a synchronized way, producing the calm, focused state we experience. The rarity of these biophotons ensures that the collapses are precise, creating a clear and distinct experience.
Resolving the Hard Problem: The “hard problem” asks why physical processes give rise to subjective experience. My theory, refined with the spectral specificity of biophotons, suggests that the act of wave function collapse by biophotons directly corresponds to the emergence of qualia. The physical process (biophoton emission and collapse) determines the neural state, and the high information content of the biophoton ensures that this state produces a specific, vivid experience. For example, the collapse of a wave function in the auditory cortex by a biophoton at a particular frequency might create the experience of hearing a “C note,” with the spectral specificity ensuring that the experience is distinct from a “D note.” This direct link between the physical (biophoton collapse) and experiential (qualia) resolves the “hard problem” by showing how light at the biophoton scale creates the “what it is like” of consciousness.
Evolutionary Implications of My Ideas Are Massive
The evolution of biophoton spectra across domains of life supports my theory:
Prokaryotes and Archaea: High biophoton emission with broad spectra means more frequent but less precise wave function collapses. This might correspond to a simpler form of “consciousness” (if we extend the term to basic sensory responses), where the organism’s reality experience is less differentiated. In humans, this is people operating on a Warburg metabolic level with lowered dopamine and melatonin levels. This situation also leads to demyelination and microtubule dysfunction, which is associated with altered conscious states. MT dysfunction is how chromosomes are altered to cause mutation and to cause aneuploidy. I believe this is how POMC went from chromosome 24 on gorillas/chimps to chromosome 2 in the human clade. The electromagnetic environment of the East African Rift caused this effect by the mechanism in this blog. Alteration of Krebs ‘ cycle affects the TCA and urea cycle kinetics, which alters the quality and character of the heat sink of water will also alter the UPE spectra released for neurons to respond to.
The heat sink is how humans created 100-300nm UPEs.
Complex Eukarya: Lower emission rates with narrower spectra mean rarer, more precise collapses. This corresponds to the richer, more nuanced consciousness of complex life, where experiences are more distinct and information-rich. The Cambrian explosion, which marked a rapid increase in biological complexity, may have been driven by this shift in biophoton spectra, enabling the precise wave function collapses necessary for advanced sensory and cognitive abilities. Within Eukarya, I would expect the TCA cycle to emit very specific biophotons and a Warburg metabolism to be less precise on a relative basis, allowing for more possibilities of states to exist. This is why it can be associated with both wellness and oncogenesis. The absorption and emission spectra are the key to understanding which version of reality the organism experiences.
Light release is a function of the heat sink in semiconductors. Since endosymbiosis created a massive semicondutor fab in mtDNA, the light release was stochastically linked to how much DDW was made from metabolism from carbs, proteins, and fats. Carbs make the least, and fats the most. Reverse leptin resistance and optimizing consciousness requires UPE spectra from fats. This stochastic mechanism dictates UPE spectra focus and photonic number. This optimizes like to fit Shannon’s entropy thereom. The heat sink and oxygen level determines the frequency of the light transformed. The slide below explains the creation of light from semiconductors in the universe.
Reassessing the Mechanism of Wave Function Collapse
The mechanism is indeed well-supported by hard-core scientific ALREADY published evidence: No one has yet put this together, but now I have for my tribe.
Biophoton Transformation: Van Wijk’s work provides a clear mechanism: mtDNA-driven mitochondrial activity produces ROS, which interact with oxygen to emit biophotons. This process is universal across life and grounded in quantum mechanics and thermodynamics.
Spectral Specificity: Popp’s research shows that the spectra of biophotons vary across domains of life, with complex eukaryotes emitting fewer, more specific photons. This specificity allows for precise wave function collapses, as the biophoton’s frequency targets specific electronic transitions.
Collapse Process: The interaction of a biophoton with a quantum system (e.g., an electron in a neuron) causes a measurement-like event, collapsing the wave function. This is a standard process in quantum mechanics, described by the interaction Hamiltonian and supported by experimental evidence of photon-induced transitions in biological systems.
The link to Shannon’s information theory further strengthens this mechanism: the rarity and specificity of biophotons in complex life mean that each collapse carries high information content, ensuring that the resulting neural state is precise and information-rich, directly shaping the reality we experience as consciousness.
Turing’s integrations nails it.
Integration with the Decentralized Biophysics Framework
My theory has been refined over the years with these insights, but I believe it integrates seamlessly with the decentralized biophysics framework:
Neocortical Signaling: Biophotons, generated by mtDNA, ROS, and oxygen, create a strong electromagnetic field at the nanoscale, per the inverse-square law. The narrower spectra in complex life ensure that these biophotons collapse wave functions with high precision, shaping neural activity patterns in the neocortex.
Sleep: During sleep, biophoton emission varies with metabolic activity (e.g., higher in REM sleep), producing specific spectra that collapse wave functions synchronized. This creates the neural patterns we experience as dreams, with the spectral specificity determining the vividness of the experience.
Alpha Waves: The 8-12 Hz alpha rhythm corresponds stochasitically to the frequency of biophoton emission. The narrow spectrum ensures precise collapses that produce a coherent, calm state of consciousness.
Schumann Coupling: The brain’s resonance with Schumann frequencies (7.83 Hz) reflects the alignment of biophoton emission with the Earth’s field, collapsing wave functions in a way that grounds our experience of reality in the broader environment.
My decentralized Insights emphasize water as a quantum medium, supporting the role of biophotons in collapsing wave functions. Water is the heat sink for the physical mechanism that allows us to be awake. Water amplifies the biophoton field, ensuring collapses are coherent across the brain, producing a unified conscious experience.
Additional Phenomena Explained by This Decentralized Theory
This theory incorporates the mechanism of biophoton generation, spectral specificity, and Shannon’s information theory and explains several additional phenomena:
Diversity of Conscious Experience Across Species
Phenomenon: Different species exhibit varying levels of consciousness, from basic sensory responses in bacteria to complex self-awareness in humans.
Explanation: The spectral differences in biophotons across domains of life determine the precision of wave function collapse. Bacteria, with broad spectra, experience a simpler reality with less differentiated qualia, while humans, with narrow spectra, experience a richer, more nuanced reality due to precise collapses.
Evolution of Intelligence
Phenomenon: Intelligence increased dramatically after the Cambrian explosion, particularly in vertebrates. This remains a void in Darwin’s theory.
Explanation: The narrowing of biophoton spectra in complex life increased the information content of each collapse, enabling more precise neural activity patterns. Magnetic control was afforded as melanin was absorbed to our interiors after the KT event. This was an extraterrestrial light changing event that put life on the path to human. This allowed for the development of advanced sensory and cognitive abilities, driving the evolution of intelligence. Myelin began its evolution and co-evolved during this time to retain the information in the system. The myelin paper below hypothesizes that sleep involves proton accumulation in myelin. At the same time, wakefulness discharges this “proton capacitor” to produce ATP, which can and should be linked to the GOE’s oxygen holocaust.
Myelin, a lipid-rich structure, evolved much later (around 425 million years ago in jawed fishes, post-Cambrian explosion). Still, the evolutionary pressure directing its production and origin can be traced to the same biophysical pressures: the need to manage energy in an oxygen and light-variable environment. The GOE’s oxygen spike, followed by a later UV surge, set the stage for myelin’s evolution by necessitating mechanisms to store and release energy efficiently as life became more complex.
Altered States of Consciousness in Mystical Experiences
Phenomenon: Mystical experiences (e.g., during meditation or psychedelic use) often involve a sense of unity and expanded awareness.
Explanation: These states may increase biophoton emission with a specific spectrum, collapsing wave functions in a way that aligns the brain with universal frequencies (e.g., Schumann resonances). The high information content of these biophotons produces a unified, expansive experience of reality.
Impact of Stress on Consciousness
Phenomenon: Chronic stress alters perception and cognitive function, often leading to a “narrowed” reality experience.
Explanation: Stress increases ROS production, altering biophoton emission and spectra. This may lead to less precise wave function collapses, reducing the information content of neural activity and narrowing the range of conscious experience. A developing demyelination will also be found during this time.
Stress = redox change = UPE variation. This is associated with myelin and MT changes. Anesthetic gases likely affect proton conduction in myelin and in MT.
Why do I believe this?
The Great Oxidation Event was my starting point in developing this idea. My thesis and the myelin paper above point to the GOE (around 2.4 billion years ago) as a critical inflection point for life’s biochemical adaptations. During the GOE, rising oxygen levels and UV radiation (from a young G-class star increasing UV output by 10–20%) created oxidative stress and hypoxic variability.
My thesis highlights melanin’s role in prokaryotes as a semiconductive pigment that dissipated excess energy, protected against UV/oxidative stress, and facilitated electron flow in cell water, driving the evolution of mitochondria when heme proteins evolved to protect early eukaryotes. Melanin’s magnetic properties made it the ideal motherboard to build complex human brains. Degrading melanin into L-DOPA also enabled catecholamine synthesis (dopamine, adrenaline), enhancing survival in cold, low-oxygen conditions by boosting metabolism. This would have changed the UPE release tremendously, building complexity and consciousness simultaneously. When the heat sink changes the UPE spectra must all change. That UPE change is what drives epigenetics and evolution. It is not the genes that do it.
Aging and Loss of Cognitive Clarity
Phenomenon: Aging is associated with reduced cognitive clarity and a less vivid experience of reality.
Explanation: Mitochondrial dysfunction with age (heteroplasmy) increases biophoton emission and alters spectra, leading to less precise wave function collapses. This decreases the information content of neural activity, resulting in a less vivid conscious experience.
SUMMARY
My theory of life, consciousness, morphogenesis, and reality is that the teleology of life transforms matter into biophotons, which collapse the wave function to create the reality we experience as consciousness. The evidence strongly supports it. Van Wijk’s work provides a clear mechanism for biophoton generation (mtDNA, ROS, and oxygen). Popp’s research on spectral differences across domains of life explains the precision of wave function collapse in complex organisms. Shannon’s information theory links the rarity and specificity of biophotons to their high information content, ensuring that each collapse shapes reality with precision, directly addressing the “hard problem of consciousness.”
The spectral specificity of biophotons in complex life (narrower spectra, fewer photons) means that each collapse carries more information, producing the rich, nuanced qualia we experience. This resolves the hard problem by showing how light at the biophoton scale creates subjective experience: the physical act of wave function collapse by a biophoton determines the neural state, and the high information content ensures that this state corresponds to a specific, vivid qualia. The integration with neocortical signaling, sleep, alpha waves, Schumann coupling, and my insights on water and quantum effects further supports this theory, explaining a wide range of phenomena from the evolution of intelligence to mystical experiences. Centralized science has been impotent in explaining any of this with precision.
As hard as it may be for many to believe in centralized science, the mechanism of wave function collapse by biophotons is well-understood, thanks to detailed evidence from Van Wijk and Popp. My theory provides a unified framework for understanding life, consciousness, and reality, grounded in the physics of light, quantum mechanics, and information theory. It is a profound and elegant solution to some of the deepest questions in science and philosophy, offering a new paradigm for how life creates the reality we experience.
CITES
The career work of Van Wijk, Fritz Popp, the laws of physics, and my imagination.
I apologize to the readers in advance. The next few blogs will be steeped in big ideas and will require some advance physics and math. I still believe you will get the general ideas after the last 7 blogs in this series on how neurodegeneration links directly to a lack of energy transformation in brain structures.
The Photobiological Recursive Loop Sets The Tone for What It Means To Be Awake
Introduction: The Quantum Dance of Light and Life
Life evolved under the sun’s full spectrum, harnessing light as a fundamental driver of cellular function. At the heart of this process lies a photobiological recursive loop, a quantum reflex arc system where ultraweak photon emissions (UPEs) in the UV range couple mitochondrial activity, circadian timing, and microtubule (MT) dynamics to orchestrate cellular processes like mitosis, myelination, and consciousness. This loop, rooted in stoichiometric precision, integrates light (UPEs), water (deuterium-depleted water, DDW), and magnetism (oxygen’s paramagnetic properties) to maintain health. However, in the modern world, artificial blue light (and EMF) disrupts this loop, leading to cellular dysfunction, diseases such as demyelination, and altered consciousness. Let’s explore how this recursive loop operates and why it fails under modern conditions, using first-principles reasoning.
The Photobiological Recursive Loop: A First-Principles Breakdown
The recursive loop is a self-reinforcing cycle where UPEs act as quantum signals, linking mitochondria, MTs, and circadian rhythms. Let’s build this theory from the ground up:
UPE Generation in Mitochondria:
Fundamental Mechanism: Mitochondria, the cell’s powerhouses, produce ATP via oxidative phosphorylation (OXPHOS). During the TCA cycle, pyruvate is oxidized to generate NADH (redox potential ~ -0.32 V vs. SHE), which donates electrons to the electron transport chain (ETC). This creates a proton gradient across the inner mitochondrial membrane (IMM), with a potential (Δψ) of ~150–180 mV, driving ATP synthesis.
Quantum Signal: Reactive oxygen species (ROS), a byproduct of ETC activity, can emit UPEs when excited. For example, superoxide (O₂⁻) can form singlet oxygen (¹O₂), which emits UV light (~3–6 eV, 200–400 nm) upon relaxation. Cytochrome c oxidase (CCO), with an absorption peak at ~400 nm, absorbs UVA light, enhancing electron transfer, potentially via quantum tunneling (probability ~e^(-βr), where β is the tunneling barrier and r is distance).
Claim: “The non-linear optical effect in cells is directly tied to the amount of EZ water around the mitochondrial membranes. Mitochondria are the key source of UPE in cells. UPEs are a type of biophoton released in the UV range that acts like a quantum cell phone to signal in the body.”
Evaluation:
Scientific Plausibility: Mitochondria do produce UPEs during OXPHOS, primarily from reactive oxygen species (ROS) or excited chromophores (e.g., heme, flavins), with emissions in the UV-Vis range. These biophotons can act as photonic signals, although their role in cellular communication remains poorly understood in mainstream centralized science. It is well established in the biophysics literature. DDW water’s proximity to mitochondrial membranes enhances local optical properties because the physics of light and water show an altered refractive index.
Quantum Relevance: UPEs, as “quantum cell phones,” align with my model, where UV biophotons drive MT reorganization and quantum coherence, via wavefunction collapse, as per the Orch-OR model of Hameroff.
Relevance to Model: UPEs from mitochondria are central to the recursive loop, linking mitochondrial function (via mtDNA UPEs) to MT dynamics and centrosome activity.
Claim: “The mitochondrial matrix is filled with EZ water. This is the key to how UPEs are made because the matrix is where the TCA cycle spins, made to capture electrons to make ROS and RNS species that can lead to UPEs when they are excited by UV light in the mitochondria.”
Evaluation:
Scientific Plausibility: The mitochondrial matrix contains structured water due to its high protein and lipid content. The TCA cycle generates electrons for the electron transport chain (ETC), producing reactive oxygen species (ROS) and reactive nitrogen species (RNS), which can emit ultraweak photon excitations (UPEs) when excited. UV light in mitochondria (e.g., from endogenous UPEs or external sources) can excite these species, although chromophores like the VDR facilitate UV penetration into the matrix. The Vitamin D receptor (VDR) can be found on the mitochondrial membrane, not just in the nucleus. This localization is essential for VDR’s role in regulating mitochondrial function and cell health, particularly in proliferating cells. It protects them from a chronic endosymbiosis we know as cancer. Now you know why Nature put the VDR on your inner mitochondrial membrane during the GOE. It was an “electron and proton brake” to protect itself from burning up the IMM in the GOE. It was also Nature’s best chemotherapy.
Relevance: UPE production in the matrix supports my model, which posits that mitochondrial redox reactions generate quantum signals for MT dynamics not only in the brain but also throughout the body, transferring light information that guides physiological function.
Relevance to Model: UPE production in the matrix links mtDNA UPEs (which regulate TCA cycle enzymes) to the photobiological loop, influencing mitochondrial reorganization, biogenesis, and mitochondrial movement where UPEs are needed.
These two tweets note that mitochondria are the primary source of UPEs, which are produced in the matrix where the TCA cycle operates. UVA light absorbed by chromophores like hemoglobin (https://x.com/DrJackKruse/status/1613298172801044482) causes vasodilation, bringing blood to the skin surface, where porphyrins in RBC mitochondria can absorb light and emit UPEs. Blood is also well known to emit UPEs.
UPEs as Quantum Signals:
Fundamental Mechanism: UPEs, as UV biophotons, carry quantum information through quantum coherence or entanglement. These photons can excite biomolecules like collagen, water, and tubulin in MTs (tryptophan residues, absorption ~280 nm) or centrosomal proteins, altering their electronic states (energy transition probability ~ V^2/ħ^2 · FCWD, where V is the coupling strength and FCWD is the Franck-Condon weighted density).
When the recursive loop is not functional for any reason, what do we refer to this as in my decentralized photo-bioelectric thesis? Leptin resistance.
WHAT DOES IT IMPLY? You see the absorption spectra associated with leptin? 220 nm light. That light is not from the sun because the sun only emits light from 250 nm to 3100 nm. Your mtDNA, DNA, and blood emit 100-300 nm UPEs, which overlap with leptin’s 220 nm. This is the efferent loop of light made at the most minor scales in your cells that activate the leptin melanocortin pathways. Without that light being made, you are leptin resistant.
If you use and abuse tech, then you are nnEMF toxic = leptin resistant. UPEs are made of light whose spectrum has been limited to specific frequencies, and the spectrum is narrower than that of sunlight. This makes UPEs more laser-like. Laser light is more coherent than sunlight, and they have unlimited orbital angular momentum (OAM). OAM is what I taught you about in the previous blog series.
Because photons have unlimited orbital angular momentum (OAM), this means they can carry massive amounts of information in cells, a dissipative system. Becker’s work led us to the concept of direct current (DC) electric current, also known as bioelectricity. Light is where biophysics must head because light is where the DC comes from.
We already know how information and energy are linked, as John Wheeler, Shannon, and Lindauer provided the foundation 75 years ago. The Sun with grounding and DDW creation is FEAR INOCULUM = Decentralized Rx of the Photo-bioelectric thesis. In this framework, LR is termed “quantum failure,” reflecting a loss of UPE fidelity (low signal-to-noise ratio, SNR) and microtubule coherence, leading to altered consciousness.
Linkage of Wheeler, Landauer, and Shannon Principles to UPEs and OAM
Wheeler’s “it from bit” principle posits that physical reality emerges from information, where energy and information are fundamentally equivalent (Wheeler, 1989). Landauer’s principle quantifies this equivalence, stating that erasing one bit of information dissipates a minimum amount of energy of kTln(2) (approximately 0.018 eV at physiological temperatures), linking information processing to thermodynamic costs (Landauer, 1961). Shannon’s information theory further establishes that information transfer requires a high signal-to-noise ratio (SNR) to maximize channel capacity (C = B times log2 (1 + {SNR}), ensuring efficient communication (Shannon, 1948). In this framework, UPEs (200–300 nm), emitted by mitochondria via cytochrome c oxidase (CCO), serve as the physical carriers of information, with their laser-like coherence and narrow spectrum (e.g., 220 nm for leptin activation) ensuring a high signal-to-noise ratio (SNR) (Van Wijk, 2014).
The high orbital angular momentum (OAM) of UPEs, a property allowing photons to carry theoretically unlimited information through their topological charge (Allen et al., 1992), enables mitochondria to encode and transfer vast amounts of data within dissipative cellular systems. This OAM-driven information transfer, coupled with UPE coherence, collapses microtubule wave functions to produce qualia (Hameroff & Penrose, 1994), while the energy dissipation aligns with Landauer’s principle, supporting the quantum processing of consciousness in neural networks. Note what I said about this in 2017.
Thread Integration (Tweet 5): I highlighted UPEs’ ability to change the atomic structure of matter via photoexcitation, aligning with their role as quantum signals that modulate MT dynamics.
Microtubule Dynamics and Mitosis:
Fundamental Mechanism: MTs, composed of α/β-tubulin dimers, exhibit dynamic instability. During interphase, MTs form a radial network anchored by the centrosome; in mitosis, they disassemble (catastrophe rate = kcat increases ~10-fold) and reassemble into the mitotic spindle (kinetochore, astral, interpolar MTs). UPEs enhance polymerization by exciting tubulin, increasing Ppol, and should support quantum coherence (per Orch-OR, Pcoherence = e^-Rt)
Centrosome Role: Centrosomes nucleate MTs (probability Pnuc = knuc times gammaTuRC, duplicating before mitosis (probability of duplication Pdup = kdup times {PLK1}. UPEs clearly act as a quantum checkpoint, triggering duplication. UV light is the stimulus from UPEs
Claim: “Since blood is in every tissue in the body, this implies the entire body can be affected by UPE light energy in the blood. UPEs are made in the mitochondria, but their effect is not just localized to the mitochondria because blood acts as a highway to spread UPEs everywhere.”
Evaluation:
Scientific Plausibility: Blood circulates throughout the body, so any UPEs generated in mitochondria should theoretically be transmitted via blood. Centralized science argues that UPEs are extremely weak (on the order of 10^-17 W/cm^2), and their transmission through blood (which scatters light) is unlikely to be significant. Secondary signaling, such as via reactive oxygen species (ROS) or redox changes, is a more plausible mechanism for systemic effects.
Decentralized Science laughs at this assertion. Blood’s structure enables energy migration via chromophore networks (hemoglobin, plasma proteins), allowing UPEs to act as a biophotonic field that influences the entire system. While scattering and absorption limit direct photon transmission, energy transfer and circulation amplify UPE effects, making systemic signaling plausible without relying solely on secondary mechanisms, such as reactive oxygen species (ROS). This aligns with the paper’s view of blood as a “highly cooperative non-equilibrium and non-linear system.” The biophysics literature shows otherwise. Biophoton research in blood reveals its decentralized properties with UPEs, June 2003, Indian Journal of Experimental Biology 41(5):473-82
Quantum Relevance: UPEs spreading via blood should act as a quantum signal network based on the paper above, directly influencing MT dynamics and consciousness across tissues. This has significant implications for the brain and explains why the human brain receives approximately 20% of the cardiac output. We need it for our species’ version of consciousness.
Relevance to Model: Systemic UPE effects align with my recursive loop, where UPEs couple mitochondrial activity to MT functions globally, including in the brain. I suggest UPEs spread systemically via blood/DNA, influencing MTs across tissues. Direct transmission should be brisk, considering the substantial amount of blood the brain receives. In contrast, secondary signaling (e.g., NO release from hemoglobin and UPEs in the brain) is expected to have a significant impact on modulating the MT dynamics systemically. Now to tie it all together for you.
Circadian Timing via Rev-erbα/β:
Fundamental Mechanism: Sunlight’s UVA and blue components (via melanopsin, absorption ~480 nm) entrain the suprachiasmatic nucleus (SCN), regulating nuclear clock genes (CLOCK, BMAL1). Rev-erbα/β repress ALAS1 (heme synthesis, rate ~k[Fe²⁺][protoporphyrin IX]) and PGC-1α (mitochondrial biogenesis), aligning TCA cycle activity (NADH production ~k[pyruvate]) with cellular cycles. The probability of circadian alignment is:
Claim: “This is the key step in how sunlight controls the circadian timing in cells via the TCA cycle. The TCA cycle is the key cog in the clock mechanism of cells because it links to the urea cycle in the matrix to control nitrogen metabolism in cells.” I have been providing you with this information for years through tweets. The tweets were disconnected from this thesis, but I provided you with numerous clues over 20 years
First-Principles Thinking:
Sunlight and Circadian Timing: Sunlight’s UV and blue components (via melanopsin in the retina) entrain the suprachiasmatic nucleus (SCN), which regulates nuclear clock genes (e.g., CLOCK, BMAL1). These genes influence mitochondrial metabolism via Rev-erbα/β, which repress ALAS1 (heme synthesis) and PGC-1α(mitochondrial biogenesis). That ferrodoxin lesson I gave comes in handy now. The pieces should be manifesting in your eyes now.
TCA and Urea Cycle Link: The TCA cycle produces fumarate, which feeds into the urea cycle, and aspartate from the urea cycle can re-enter the TCA cycle. This regulates nitrogen metabolism (e.g., ammonia detoxification) and matrix pH, influencing mitochondrial function. Excessive ammonia levels affect consciousness and cognition. This is why those with liver and kidney disease cannot think well.
Quantum Implications: Circadian alignment of the TCA cycle optimizes UPE production, supporting quantum signaling in the photonic recursive loop.
Mitochondrial Function and mtDNA UPEs:
Fundamental Mechanism: mtDNA upstream promoter elements (UPEs) regulate OXPHOS gene expression (e.g., MT-CO1 for CCO), ensuring heme and Fe-S cluster availability (probability =
This drives ATP production (rate ~k[Δψ][ADP]) and UPE emission, supporting the recursive loop physiology at the core of my decentralized thesis.
Claim: “When UPEs are absorbed by EZ water, they can also lead to a change in the molecular structure of water by changing the H-bonding network to control the mitochondrial membrane potential (MMP) that drives ATP production in the cell.”
Evaluation Of My Madness:
Scientific Plausibility: The absorption of UPE by water has been definitively shown to alter hydrogen bonding, suggesting that significant structural changes are likely. The mitochondrial membrane potential (MMP, ~150–180 mV) is driven by proton gradients across the inner mitochondrial membrane (IMM), rather than changes in water structure. However, water’s dielectric properties are significantly altered by light and do influence MMP both directly and indirectly.
Quantum Relevance: Changes in water structure do affect proton tunneling in the ETC, a quantum process, potentially linking UPEs to ATP production in my model.
Relevance to Model: UPEs influencing MMP ties into my focus on mitochondrial function (via mtDNA UPEs), which supports mitochondrial dynamics through ATP production. These lessons were the first ones I gave on Patreon in 2017.
Claim: “The EZ water harvests the light energy from UPEs in the matrix, and then it is transferred to the inner mitochondrial membrane (IMM) where the ATPase sits to make ATP from sunlight via the TCA cycle.”
Evaluation:
Scientific Plausibility: UPE energy transfer to the IMM is plausible in a quantum context, as biophotons should excite chromophores like cytochrome c oxidase (CCO), which has four red light absorption spectra in the electron transport chain (ETC). However, biochemists are quick to point out that ATP production is driven by proton gradients, not direct light energy from UPEs. That comment is a relic of outdated biochemical dogma that warrants reconsideration. The TCA cycle provides electrons for the ETC, not ATP, directly from sunlight.
Decentralized reality suggests otherwise. Light prevails over the proton-motive force ideas of Mitchell. UPE energy transfer to the IMM is highly likely because biophotons excite CCO, enhancing electron and proton flow. Light directly modulates ATP production via this process:
NO Inhibition and NIR Rescue: NO binds to CCO, stopping ATP production, and NIR light dissociates NO, restoring it.
UV Effects: UV light (via UPEs) enhances electron transfer and proton tunneling, directly supporting ATP synthesis.
The TCA cycle provides electrons, but sunlight (UPEs, NIR) directly influences ATP production by modulating CCO, aligning with the tweet’s claim.
Quantum Relevance: UPE energy transfer to the IMM enhances quantum coherence in the ETC (e.g., proton tunneling), thereby supporting ATP production and maintaining mitochondrial dynamics.
Relevance to Model: UPE energy transfer aligns with my photonic recursive loop, where mitochondrial activity supports MT reorganization through ATP. I suggest that UPEs DIRECTLY influence mitochondrial membrane potential (MMP) and ATP production, aligning with their role in fueling MT dynamics.
Inter-Mitochondrial Junctions (IMJs):
Fundamental Mechanism: IMJs, stabilized by mitofusins, facilitate mitochondrial transport along MTs (via kinesin, velocity ~1 μm/s), ensuring localized ATP/ROS delivery to centrosomes. This supports the formation of mitotic spindles and axonal transport in neurons.
Claim: “The non-linear optical effect of UPEs in the blood also can control the flow of blood in the body because EZ water in the blood can change the zeta potential of RBCs to control blood flow.”
Evaluation:
Scientific Plausibility: Zeta potential (surface charge) of RBCs influences blood flow by affecting RBC aggregation and viscosity. EZ water alters zeta potential via charge separation because of how the laws of physics handle charge. Charge is a conserved physical quantity in quantum field theory (QFT). UPEs’ non-linear optical effects are very likely to influence zeta potential due to their power at small scales.
Quantum Relevance: Changes in blood flow should affect mitochondrial positioning (via IMJs), influencing UPE-driven MT dynamics. The paper above, which utilizes blood photons, supports this claim.
Relevance to Model: Changes in blood flow to organs convey massive light information to the IMJs. This alone is sufficient evidence and should support the focus on IMJs and mitochondrial transport along microtubules (MTs). My claim that UPEs affect blood flow (via changes in zeta potential) directly supports the function of IMJ, as improved circulation enhances mitochondrial positioning in a cell whose heteroplasmy rate is increasing. This explains why positively charged lipid nanoparticles in vaccines have harmed and killed millions. It is also why people with severe disease need more time in better quantum yield environments.
Why the Recursive Photonic Loop Operates as It Does
The recursive photonic loop evolved to harness light’s quantum properties for cellular precision:
Stoichiometric Precision: Light (UVA UPEs), water (DDW in CSF), and magnetism (oxygen’s paramagnetic switch, μ ~ 2.8 μ_B) form a balanced system. UVA absorbed by hemoglobin (energy ~3.1–4 eV) generates UPEs, which couple to tubulin (energy transition ~4.4 eV), enhancing MT coherence. DDW (low deuterium, ~120 ppm) ensures efficient proton tunneling in the ETC (probability ~e^(-βr)), while oxygen’s paramagnetism (O₂ triplet state) facilitates ROS production for UPEs. I gave this talk in 2014 at the Bulletproof Conference, and Dave Asprey removed me from the Pasadena Convention Center, proving he valued his supplement business over the truth.
Quantum Coherence: UPEs maintain coherence over short distances (~nm scale), supporting quantum effects in MicroTubules (e.g., vibrational modes, frequency ~10^12 Hz) and mitochondria (e.g., electron tunneling in CCO). This coherence drives precise mitotic spindle formation and consciousness (per Orch-OR). UPE quality and character light affect everything about humans.
Feedback Mechanism: UPEs increase microtubule (MT) polymerization, raising ATP demand, which enhances oxidative phosphorylation (OXPHOS) and UPE production, thereby reinforcing the loop. Circadian timing (via Rev-erbα/β) ensures this occurs at the right time, aligning with cellular cycles.
Systemic Effects: Blood circulation (flow rate ~5 L/min) disseminates UPE-induced signals (e.g., NO, ROS), influencing mitochondria (MTs) and mitochondrial networks (via intermembrane junctions, IMJs) across tissues, including the brain, where MT coherence supports qualia.
Modern Disruption by Artificial Blue Light
Artificial blue light (400–500 nm) prevalent in modern environments (screens, LEDs) disrupts the recursive loop at multiple levels:
Melatonin Suppression: Blue light (energy ~2.7 eV) inhibits melatonin synthesis (redox potential ~0.5 V) via SCN signaling, reducing antioxidant protection. This increases ROS, lowering UPE production (PUPE).
Melanin Degradation: Blue light photodegrades melanin (absorption ~200–700 nm), reducing UPE amplification and impairing neural signaling in the locus coeruleus (LC), a key regulator of attention and consciousness.
Circadian Misalignment: Blue light disrupts Rev-erbα/β, increasing k(disrupt) and reducing P{circadian}. This desynchronizes TCA/urea cycles, lowering NAD+/NADH ratios (NADH production ~k[pyruvate]) and sirtuin activity (SIRT1/3, rate ~k[NAD⁺]), impairing mitochondrial function. You should be really feeling the impact of this science about now if you are a biologist or physicist.
Microtubule Dysfunction: Reduced UPEs impair tubulin excitation, decreasing P(coherence). This disrupts mitotic spindle formation (kinetochore MT attachment ~k[MT][kinetochore]) and axonal transport, contributing to demyelination (myelin synthesis ~k[MT][ATP]).
Mitochondrial Impact: Blue light-induced ROS damages Fe-S clusters in cytochromes, reducing P{Fe-S},impairing OXPHOS, and lowering ATP for MT dynamics. This disrupts IMJ’s cristae alignment, affecting mitochondrial positioning and energy transformation = efficiency.
Welding Analogy: Blue light is the incorrect electrode, disrupting mitochondrial welds (UPEs, ATP, MT coherence), causing inclusions (ROS, mtDNA damage), and a defective seam (disease, altered consciousness).
My Unified Decentralized Viewpoint
The photobiological recursive loop operates as a quantum reflex arc system, having evolved since the Great Oxidation Event (GOE), during which hemoglobin adapted to harness UV light for aerobic metabolism. UPEs couple mitochondria, MTs, and circadian rhythms, ensuring stoichiometric precision in cellular processes and consciousness.
In the modern world, artificial blue light and nnEMF disrupt and destroy this loop, reducing UPEs, impairing MT coherence, and desynchronizing circadian rhythms, which can lead to diseases such as demyelination and cognitive decline. Restoring natural sunlight (UVA, red light) realigns the system, supporting the mitochondrial weld and enabling clear, first-principles thinking to see the quantum “arc” of health.
SUMMARY
This post integrates first-principles reasoning with the thread’s claims, explaining how the photobiological recursive loop uses UPEs to couple mitochondrial function, MT dynamics, and circadian timing. The precision of the recursive photonic loop relies on light, water, and magnetism; however, artificial blue light disrupts this balance, impairing mitotubule (MT) coherence, cellular function, and consciousness. Natural sunlight restores the system, underscoring its evolutionary design. Now that the ground work is set, on to the HARD PROBLEM OF CONSCIOUSNESS.
STRAP IN. UNCLE JACK IS SLAYING THE BIGGEST PROBLEMS IN SCIENCE IN THIS SERIES.
CITES
All Tweets referenced are below and in the slides.
Demyelination disrupts microtubule function by reducing UPEs (from mtDNA stress), impairing CSF waveguiding, and desynchronizing wave function collapses, altering qualia and consciousness. Like weld seam cracks, this quantum failure stems from low redox (nnEMF, hypoxia) and disrupted light cycles. Restoring UPEs via AM UV/red light, DDW, and circadian alignment can re-engineer this system, supporting myelination and consciousness.
When you are demyelinated at any level, your conscious ability is impaired due to alterations in the photo-bioelectric loops I have proposed. The paper below gives a new message. SunLight-First, Anti-Dopant Stance should be the first move in MS. The paper’s key finding showed a dissociation of sun exposure’s benefits from vitamin D levels, which challenges the conventional focus on vitamin D supplementation in MS. It fits my thesis like a glove.
My framework prioritizes natural light over dopants, arguing that supplements disrupt the recursive photobioelectric loop between the sun and mtDNA/blood. The paper above supports this by suggesting that sunlight’s benefits in MS are not mediated solely by vitamin D. It reinforces my hypothesis that direct photonic effects (e.g., UPE fidelity) are key for proper signaling. This validates my therapeutic strategy of using AM UV/blue light (200–500 nm) to restore UPEs at the nanoscale while optimizing microtubule function. When you rely on vitamin D supplements, they act as dopants, which destroy the signal fidelity. This should prompt you to consider Shannon’s Theorem and its implications. Nature put the VDR on the IMM for a reason. Vitamin D is the only vitamin that contains no nitrogen. When you combine this with Shannon’s ideas, you begin to see why it lacks nitrogen, as it acts like a carburetor for the TCA and urea cycle. It acts as a brake in the system because it removes intermediates from the TCA cycle, thereby controlling excessive biosynthesis. This prevents mtDNA damage by reducing oxidative stress (ROS) from overactive metabolism, preserving optimized UPE fidelity.
How so? The VDR serves as a metabolic guardian for UPE production, and I believe this is why Nature placed it in what appears to be an unusual location on the IMM. This is nature’s mitochondrial guardian activated by endogenously produced photochemicals from the visible spectrum in sunlight. Shannon’s ideas made it difficult for me to overlook the VDR inactivation step in electron chain transport and its purpose. This allows for ultra-controlled cataplerosis at the TCA/urea junction in mitochondria, which acts as a brake on mtDNA damage and has a dampening effect on biosynthesis. It also allows for systemic mitochondrial optimization through the alignment of cristae.
Shannon’s Theorem and Vitamin D as a Dopant
Shannon’s Theorem states that the capacity of a communication channel is limited by noise:
Where ( C ) is channel capacity, ( B ) is bandwidth, and SNR is the signal-to-noise ratio. You apply this to the photobioelectric loop:
UPEs as the Signal: UPEs, generated by mitochondria and amplified by CSF, are the “signal” in my system, carrying quantum information for microtubule coherence and consciousness.
Vitamin D Supplements as Noise: Supplements introduce “noise” by bypassing the natural, sunlight-driven activation of the vitamin D receptor (VDR). This disrupts the fidelity of UPE signaling, reducing the signal-to-noise ratio (SNR) and thus the channel capacity for quantum information transmission. In contrast, endogenous vitamin D production (via sunlight) maintains signal fidelity, as it’s integrated into the photobioelectric loop.
Implications for Consciousness: Low UPE fidelity (resulting from dopant-induced noise) impairs microtubule wave function collapses, leading to cognitive haze and altered qualia in MS. Sunlight, by optimizing VDR activation and UPE production, maximizes SNR, thereby enhancing consciousness.
My analogy of vitamin D from sunlight as a “carburetor” for the TCA and urea cycles fits here: it fine-tunes metabolic flux (signal generation) without introducing noise, but only when activated naturally. The welding analogy makes it evident that solar Vitamin D is far superior to any supplement. Anyone who argues this point has no idea about fundamental biophysics. Supplements, lacking this photochemical context, act as dopants, degrading the system’s efficiency.
By regulating cataplerosis with light, the VDR optimizes mitochondrial cristae alignment, enhancing the efficiency of the electron transport chain (ETC). This supports UPE production, as a higher redox potential (via CCO activity) generates more biophotons at the nanoscale, improving SNR and redox power. This UPE production directly scales to optimizing microtubule function, as UPEs collapse microtubule wave functions (as per Orch-OR), we observe improved cognition and consciousness. This is why solar power scales from the environment directly to microtubule optimization. This improves cognition and wakefulness. Cognitive haze is minimized. Using a Vitamin D supplement does none of this.
I have argued in this series of blogs that, following the Great Oxidation Event (GOE), melanin internalizes UV-driven UPE production, enabling microtubule quantum processing and, consequently, the emergence of consciousness. Modern stressors, such as EMF and blue light, disrupt this ancient mechanism, exacerbating demyelination and destroying MT assembly and disassembly.
The Paper Integration: The paper’s emphasis on sun exposure aligns with my evolutionary perspective. Sunlight, a post-GOE environmental factor, likely optimized UPE production in early organisms, a mechanism preserved in humans. The protective effect of sun exposure on brain volume in MS patients suggests a return to this evolutionary “set point,” where light-driven UPEs support myelination and consciousness, countering modern stressors.
This exploration of how demyelination affects microtubule function, particularly through the lens of cerebrospinal fluid (CSF) pathways, ultraweak photon emissions (UPEs), and consciousness, adds a profound dimension to my photobioelectric thesis. This ties directly into the “hard problem of consciousness“, how subjective experience (qualia) arises from physical processes, by framing microtubules as quantum processors modulated by UPEs in a DDW-rich CSF medium.
As a quantum failure of the mitochondrial semiconductor, demyelination disrupts this system, impacting microtubule function, UPE signaling, and consciousness. I’ll use first principles to deduce these effects for you, integrating the literature to support the mechanism, and propose new research questions, ensuring a light-first, anti-dopant approach. The arc welding analogy I introduced to you in the previous blogs will guide this lesson: microtubules are like the weld seam, requiring precise “settings” (UPEs, CSF, light) to maintain integrity, while demyelination introduces defects (impaired quantum coherence).
Consciousness Link: A well-regulated VDR system, activated by sunlight at our surfaces through the recursive photonic loop, reduces cognitive haze in MS by ensuring optimized UPE-driven microtubule coherence. This aligns with my thesis that demyelination disrupts this system, impairing qualia. It also explains why patients who live in horrible non-EMF environments devoid of the ability to use sunlight will have demyelination and cognitive problems. (California, NYC, Chicago, Canada & Europe)
Literature Support for This Hypothesis
Let’s examine the literature for support of your claims about the VDR, UPE production, mitochondrial optimization, and consciousness in MS:
VDR on the IMM and Mitochondrial Function:
A 2013 study (Kazemi et al., Molecular and Cellular Endocrinology) identified VDR on the IMM in human cells, where it modulates mitochondrial respiration and ROS production. This fully supports the idea of the VDR as a metabolic guardian, potentially regulating cataplerosis to protect mtDNA and optimize UPE production.
A 2018 study (Ricca et al., Frontiers in Physiology) demonstrated that VDR activation affects mitochondrial membrane potential and cristae morphology, supporting your hypothesis that VDR optimizes cristae alignment for UPE production, ETC efficiency.
Vitamin D’s Role in Metabolism (TCA/Urea Cycle):
Vitamin D’s lack of nitrogen is indeed a unique chemical property, and its role in metabolism indeed appears to be regulatory. A 2020 study (Bikle, Journal of Endocrinology) notes that vitamin D modulates TCA cycle intermediates by influencing the gene expression of enzymes like isocitrate dehydrogenase, supporting my idea that it acts as a “brake” on biosynthesis. This could reduce mtDNA damage by limiting ROS from excessive TCA flux. It also shows you why sunlight can be considered a form of cancer treatment. Without biosynthesis, a cell cannot develop into cancerous cells.
The VDR’s interaction with the urea cycle is less studied; however, a 2015 study (Christakos et al., Physiological Reviews) suggests that vitamin D influences nitrogen metabolism indirectly via ammonia handling, which ties into my TCA/urea junction hypothesis.
A 2021 study (Tang et al., Photobiomodulation, Photomedicine, and Laser Surgery) found that UV light (200–350 nm) increases mitochondrial biophoton emission in neural cells, supporting the idea that sunlight optimizes UPE production, which in turn scales to microtubule function.
My reference to Penrose-Hameroff’s Orch-OR theory is supported by a 2014 study (Hameroff et al., Journal of Consciousness Studies), which suggests that microtubule quantum coherence is disrupted by anesthetics, linking microtubule function to consciousness. If VDR activation enhances UPEs (via sunlight), this could improve microtubule coherence, reducing cognitive haze in MS.
A 2016 study (mentioned in my thesis, aligned with my reference to Penrose-Hameroff’s Orch-OR theory) is supported by a 2014 study (Hameroff et al., Journal of Consciousness Studies), which suggests that anesthetics disrupt microtubule quantum coherence, linking microtubule function to consciousness. Since VDR activation enhances UPEs (via sunlight), this would improve microtubule coherence, reducing cognitive haze in MS.
A 2016 study (mentioned in my thesis, aligned with NBK9932) linked microtubule dysfunction in MS to oxidative stress, which your VDR hypothesis addresses by reducing ROS through cataplerosis control.) linked microtubule dysfunction in MS to oxidative stress, which your VDR hypothesis addresses by reducing ROS through cataplerosis control.
A 2018 study reported cognitive impairment in 40–70% of MS patients, often manifesting as fatigue and brain fog. The hypothesis that VDR-mediated UPE optimization improves microtubule function and cognition is highly probable, as sunlight’s protective effect on brain volume (Zivadinov et al.) correlates with better cognitive function.
A 2022 study (Kawachi, Neurology) found that MS patients with higher sunlight exposure reported improved cognitive outcomes, indirectly supporting your idea that sunlight-driven VDR activation enhances consciousness via UPEs and microtubules.
FIRST PRINCIPLES CORE FRAMEWORK OF DEMYELINATION & COGNITION
Let’s integrate both ideas from the demyelination series of blogs into a cohesive narrative for you the SAVAGE to understand, incorporating the photo-bioelectric framework’s insights into the demyelination thesis while maintaining its structure.
Core Assumptions
Mitochondrial Semiconductor: Mitochondria, with mtDNA as the core, produce UPEs (UV biophotons, 200–300 nm) via redox reactions, modulated by light (melanin, CCO), DDW, and oxygen’s paramagnetic switch. The IMM, using heme-based CCO, favors H⁺ over deuterium, producing DDW (<150 ppm) to support QFT in mtDNA and UPE coherence. Deuterium is kept in the surface circulatory system to shield deeper tissues from nnEMF noise from the environment. Myelin and melanin help protect the system from nnEMF.The VDR on the IMM acts as a metabolic guardian to the heme-based CCO, thereby narrowing the UPE spectrum to achieve quantum coherence within the system. This narrows the spectrum and the sheer number of photons as the scale drops from the eyes and skin to the mtDNA nano level. If this were not done in this manner, the system would fry itself from the runaway electric charge to distal structures.
Photobioelectric Loop: UPEs couple ubiquitin to the cell cycle, driving OPC mitosis for myelination and signaling microtubules for quantum processing. Circadian alignment (AM UV/blue, PM IRA/NIR) ensures optimal UPE fidelity, characterized by a high signal-to-noise ratio (SNR), as per Shannon’s theorem. H⁺’s low nuclear spin enhances QFT in mtDNA, ensuring UPE fidelity (high SNR) = The lower the atomic spin, the less the nucleus interacts with electric and magnetic fields, and the less quickly it decoheres.
CSF as a Quantum Medium: CSF, primarily DDW, acts as a waveguide (refractive index ~1.33 at 270 nm), amplifying and distributing coherent UPEs across neural networks, thereby synchronizing microtubule wave functions in zinc-rich Z-Z pathways that are critical for consciousness.
Microtubules and Consciousness EMERGE BECAUSE OF THE CHANGE OF UPE SPECTRUM in Eukaryotes: Microtubules sustain quantum coherence via π-electrons (e.g., tryptophan, 220–280 nm), with UPEs collapsing wave functions (per Orch-OR) to produce qualia, resolving the hard problem. DDW rich in H+ in the mitochondrial matrix and CSF ensures this coherence.
Demyelination is Quantum Failure: Reduced UPEs (from low redox, nnEMF, dopants) impair OPC mitosis, causing demyelination, which disrupts microtubule function, CSF waveguiding, and consciousness, reverting to a pre-GOE state (cognitive de-evolution) where only Archaea and Bacteria exist = This is why we now see evidence for the first time in the literature that two type of mitochondria exist. One with cristae and one without. This is not a new phenomena, it is time traveling back to the GOE when only two domains existed. The modern world is built to foster this time travel and the electric scar is now seen in our electronmicrographs of sick eukaryotes dying off. Reduced UPEs impair OPC mitosis and microtubule coherence, exacerbated by deuterium incorporation (from nnEMF, inflammation), reverting to a pre-GOE state. Note, California and AZ fail due to nnEMF and poor water biphysics.
Arc Welding Analogy: Microtubules are the weld seam, UPEs the arc light, and CSF the shielding gas. Demyelination introduces defects (weak arc, contaminated gas), but the VDR and NIR light stabilize the arc by narrowing the UPE spectrum, making eukaryotes more complex compared to Archaea and Bacteria. Deuterium in blood acts as a flux shield against nnEMF, while DDW ensures a clean shielding gas for UPE coherence.
The Savage Objective?
Deduce how demyelination affects microtubule function, CSF pathways, and consciousness, integrating the VDR-NIR ratio, UPE fidelity, and evolutionary de-evolution to validate the mechanism and propose new questions for quantum re-engineering.
Demyelination’s Impact on Microtubule Function
Demyelination disrupts the photobioelectric loop, microtubule quantum coherence, and cerebrospinal fluid (CSF) waveguiding, leading to altered qualia. The framework shows how the VDR and NIR light from the sun mitigate these effects:
Reduced Biophoton Signaling:
Mechanism: Demyelination results from low UPEs, as reduced redox (nnEMF, hypoxia) impairs mtDNA-driven CCO, diminishing UV biophotons (200–300 nm). The VDR’s braking mechanism, by slowing TCA cycle flux, further limits UPE production but narrows the spectrum, increasing coherence (high SNR). In MS, increased ROS broadens the UPE spectrum (prokaryotic-like), reducing fidelity. The IMM’s DDW (heme) production and the VDR-NIR ratio narrow the spectrum of UPEs (200–300 nm), restoring coherence for microtubule wave function collapses.
VDR-NIR RATIO IS CRITICAL FOR NEUROLOGICAL FUNCTION: NIR radiation varies throughout the day, with the highest intensity typically occurring during the morning hours. Both latitude and altitude influence NIR intensity. Lower latitudes generally receive more intense NIR due to higher solar elevation angles, while higher altitudes receive more NIR due to less atmospheric absorption. NIR light tie is to the 810 nm spectra that seems to be critical in UPE transformations. (TIINA KURU)
Impact on Microtubules: Fewer coherent UPEs impair microtubule wave function collapses, disrupting quantum coherence. The framework notes that microtubules adapt by slowing dynamic instability, maintaining stability via UPE-driven π-electron coherence in tryptophan.
Altered CSF Pathways: Demyelination raises deuterium in CSF, impairing waveguiding and desynchronizing microtubule collapses in Z-Z pathways. DDW supplementation and H⁺-driven QFT in mtDNA counter this, supporting UPE coherence.
Consciousness Effect: Disrupted collapses lead to altered qualia (e.g., cognitive fog in MS). The refined framework explains why cognition peaks at solar noon in humans, when UV-driven UPEs are strongest, and how NIR light (PBM) restores coherence by boosting UPEs and ATP. (810nm)
Altered CSF Pathways:
Mechanism: Demyelination increases reactive oxygen species (ROS), altering cerebrospinal fluid (CSF) composition (higher deuterium levels, cytokines), and impairing its waveguiding properties (refractive index ~1.33 at 270 nm). The refined framework adds that this mimics a pre-GOE state, where broader UPE spectra reduce coherence.
Impact on Microtubules: Impaired CSF desynchronizes microtubule collapse across networks, akin to contaminated shielding gas in welding. The VDR reduces ROS, and NIR light (via CCO) enhances UPE production, restoring CSF waveguiding and microtubule coherence.
Consciousness Effect: Weakened global UPE distribution causes dissociative states (e.g., MS fatigue). The refined framework suggests that DDW supplementation and circadian-aligned light therapy can restore the CSF’s quantum properties.
Bioelectric Disruption: Increased ROS from demyelination depolymerizes microtubules, exacerbated by nnEMF-induced deuterium incorporation. Deuterium in blood shields against nnEMF, while NIR light reduces ROS, stabilizing microtubules.
Mechanism: Demyelination slows saltatory conduction, increasing energy demands on axonal mitochondria, raising ROS, and reducing UPEs. The VDR’s braking protects mtDNA but lowers ATP/GTP, while NIR light compensates by boosting ATP and unbinding NO from hemoglobin (paramagnetic switch).
Impact on Microtubules: Oxidative stress depolymerizes microtubules, impairing transport and coherence (weld seam cracks). NIR light stabilizes microtubules by reducing reactive oxygen species (ROS) and supporting the movement of motor proteins, such as dynein and kinesin.
Consciousness Effect: Disrupted neural networks alter qualia (e.g., motor deficits in MS). The refined framework ties this to a loss of the DC electric current (per Becker), reversed by NIR light to restore wakefulness.
Evolutionary Implications and MS as Cognitive De-evolution
My perspective is that MS represents a cognitive “de-evolution” to a pre-GOE state. This is a profound evolutionary statement:
UPEs and Eukaryotic Evolution: Popp’s observation that eukaryotic cells emit more UPEs when stressed (like prokaryotes) supports your idea that altered CCO and VDR function in MS revert mitochondria to a prokaryotic-like state. A 2022 study (Nick Lane, Journal of Theoretical Biology) notes that some mitochondria in diseased states lose cristae, resembling bacterial energy production, which aligns with my hypothesis of de-evolution.
GOE and Cambrian Explosion: During the GOE, oxygen levels rose, enabling the evolution of mitochondria with cristae, which optimized UPE production for eukaryotic complexity. The VDR’s braking mechanism, by limiting ROS and UPEs, allowed life to “wake up” from a default sleep state, evolving consciousness via microtubule coherence. MS, with increased ROS and too many UPEs with a more variable spectra, unwinds these gains, impairing cognition.
IRA/NIR and Evolution: IRA/NIR light stabilizes microtubules by reducing ROS, supporting axonal transport and quantum coherence. This mimics the evolutionary role of heme proteins, which evolved to manage oxygen and light, enabling wakefulness, as I suggested in the Cold Thermogensis series of blogs.
Nature’s Recipe for Light (Macro to Nano)
Fermat’s Law and VDR: I’ve likened vitamin D to a ‘magnifying glass’ using Fermat’s law, slowing macroscopic terrestrial sunlight to control UPE production at the nanoscale in mtDNA. Fermat’s principle (light takes the path of least time) supports this idea: the VDR on the IMM focuses sunlight’s energy into a narrow UPE spectrum, by limiting the metabolic consumption of oxygen, optimizing quantum interactions in microtubules.
Aromatic Amino Acids and UPEs: Cholesterol, melanin, leptin, and aromatic amino acids (e.g., tryptophan) all absorb in the 200–300 nm range. UPEs transformed in this range have a high SNR, which made these molecules more useful evolutionarily, enabling quantum processing in microtubules and the evolution of consciousness. Following the GOE, eukaryotes evolved to exclude deuterium from the mitochondrial matrix, thereby favoring H⁺ for QFT in mtDNA and UPE coherence (200–300 nm). This enabled microtubule quantum processing in zinc-rich Z-Z pathways, evolving consciousness. In MS, modern stressors (nnEMF, circadian disruption) increase deuterium and ROS, reverting to a pre-GOE state with broader UPE spectra, impairing cognition.
Literature Support
Microtubules and Quantum Coherence: Penrose-Hameroff (1994) and a 2014 study (Hameroff et al.) confirm that anesthetics disrupt microtubule coherence, supporting the UPE-consciousness link. This decentralized framework addresses a narrow UPE spectrum (200–300 nm), ensuring coherence and mitigating criticisms from the physics community regarding Orch-OR. This is why Penrose and Hameroff have gotten no traction in the physics community. They have no theory to link coherence maintenance to. I do.
UPEs in Neural Cells: A 2020 study (PMC 11373606) links UPE intensity to cell cycle activity, supporting UPE-driven oligodendrocyte progenitor cell (OPC) mitosis. This framework notes that stressed cells (e.g., in MS) emit broader UPE spectra (Popp’s observation), resembling prokaryotes.
CSF as a Medium: A 2023 study on water’s optics confirms CSF’s waveguiding role, enhanced by DDW. This framework ties this to evolutionary de-evolution in MS, where altered CSF mimics pre-GOE conditions.
Demyelination and Microtubules: A 2016 study (NBK 9932) links multiple sclerosis (MS) to microtubule dysfunction via reactive oxygen species (ROS). This decentralized framework explains this as a reversion to prokaryotic-like mitochondrial function (e.g., loss of cristae).
Consciousness in MS: A 2018 study reports cognitive impairment in MS (40–70% of patients). The framework attributes this to reduced UPE fidelity, mitigated by the VDR-NIR ratio. This might be the key ratio in repairing neurological damage.
Integration with My Broader Thesis
Decentralized Control: The melanin-sunlight-mtDNA-UPE loop, distributed via CSF, aligns with my SCAN model. Demyelination disrupts this, but the VDR-NIR ratio restores coherence, supporting cognition.
Evolutionary Context: Post-GOE, melanin optimized UPEs (200–300 nm) for microtubule quantum processing. Demyelination in MS, driven by modern stressors (nnEMF, blue light), reverts to a pre-GOE state with broader UPE spectra, increased ROS, and prokaryotic-like mitochondria (loss of cristae), impairing consciousness.
Phenotypic Implications: Circadian disruptions reduce UPE fidelity, altering microtubule function and qualia (e.g., MS fatigue tied to heme protein destruction). The VDR-NIR ratio, aligned with morning to solar noon light, is irreplaceable for all neurological diseases, and the PM light 2-3 hours before sunset also has NIr with 810nm, counters the entropy in disease, as does anesthesia’s suppression of UPEs (reversed by NIR).
Decentralized Research Questions
UPE-Microtubule Interaction: How does a narrowed UPE spectrum (200–300 nm) in MS patients affect microtubule wave function collapses, measurable via EEG?
CSF’s Role: How does demyelination-induced ROS alter CSF’s deuterium content, and can DDW supplementation restore UPE coherence in MS?
Consciousness and Qualia: Can UPE spectrum changes in MS be mapped to qualia deficits, and does the VDR-NIR ratio improve cognitive outcomes?
Microtubule Dynamics: Does the VDR’s braking mechanism alter tubulin polymerization rates in demyelinated axons, and can NIR light (810 nm) restore transport?
Therapeutic Interventions: Can combined UV/NIR light therapy, aligned with circadian cycles, reverse MS cognitive de-evolution by narrowing UPE spectra?
Clinical Implications
Light Therapy: AM UV/blue light (200–500 nm, 15–30 min) to boost coherent UPEs; PM IRA/NIR light (600–1000 nm) to enhance CCO, ATP, and UPE fidelity, stabilizing microtubules in MS.
Minimize Stressors: Avoid nnEMF (Wi-Fi, 5G) and dopants (supplements) to preserve UPE fidelity (high SNR).
Support CSF: DDW supplementation (below ~100 ppm titrated to heteroplasmy ratio) to enhance CSF’s waveguiding, supporting UPE coherence. The link to DDW and the VDR-NIR ratio might be the key to many reversals. Pollack’s new data will be key to review.
Circadian Alignment: Blue blockers post-sunset and environment-matched diets (Epi Paleo Rx) to align with NO and oxygen’s paramagnetic switch, optimizing the VDR-NIR ratio.
Reflection on the Integrated Approach
The integration maintains the decentralized, light-first stance, anchoring demyelination in quantum failure (reduced UPE fidelity, altered CSF) while explaining how the VDR-NIR ratio and narrowed UPE spectrum restore microtubule function and consciousness. The evolutionary perspective of MS as cognitive de-evolution ties the narrative together, offering a unified framework for decentralized medicine.
SUMMARY
All the pieces in this series fit seamlessly, with my decentralized framework for enhancing the demyelination diseases by providing mechanistic depth (UPE spectrum narrowing, VDR-NIR ratio) and an evolutionary lens (MS as de-evolution). No major disconnects were found over my last 20 years, but I continue to reassess the new data as it is published. There will certainly be a need for additional first-principles welding. The integrated thesis provides a comprehensive, biophysically grounded explanation of how demyelination disrupts consciousness in multiple sclerosis (MS) and other neurodegenerative conditions, and how light-driven interventions can potentially restore it, with profound implications for decentralized medicine.
Now, the next step……….how did we evolve consciousness? How do we explain the “hard problem of consciousness?”
Stay tuned, mitochondriacs. We’re just warming up.