The Great Oxygenation Event (~2.4 Ga) transformed Earth’s atmosphere, increasing electrical conductance through water vapor, sulfate aerosols, and reactive species like NOx. Lightning and UV-driven ROS pressured archaea and bacteria into endosymbiosis, where an oxygen-respiring bacterium became the proto-mitochondrion.
Mitochondria evolved as cellular magnetospheres, using DHA, heme proteins, metal SODs, and proton gradients to manage electron flow, enabling eukaryotic complexity and later innovations like myelin. Artificial blue light and nnEMF disrupt mitochondrial function today, increasing conductance and ROS in a fractal echo of GOE dynamics, driving chronic disease. Decentralized medicine must restore biophysical harmony by addressing light, water, and magnetism to make sense of chronic diseases. Our recent electromagnetic exposures fully mirror the evolutionary conditions that birthed eukaryotes.
Clarified Evolutionary Timeline: Endosymbiosis (2 Ga) predates the Cambrian explosion (540 Ma). The latter reflects oxygen-driven multicellularity, not the initial eukaryotic event. This timeline comprehension is critical in understanding why we must emphasize GOE as the endosymbiosis trigger and Cambrian as a downstream effect. The KT event was another downstream effect, particularly a light-stress event for mammals.
Strengthened Conductance Model: Incorporated aerosols, trace gases, and water vapor were the conductance drivers, and this is grounded in modern atmospheric science (e.g., volcanic lightning studies). Geologists have added BIFs and stromatolites as geological evidence of GOE’s impact that has scarred living systems.
Nature’s Fractal Analogy: The magnetosphere-IMM parallel is robust idea clarified as charge-separation system. We needed to add biophysics (e.g., DHA’s π-electrons, structured water) to bridge planetary and cellular scales to see how this sculpted the IMM.
Modern Relevance To Light Stress as the Modern Cheisel of Disease: Biophysics links blue light/nnEMF to specific mitochondrial targets (CCO, mtDNA, SOD) and chronic disease, aligning with my decentralized photo-bioelectrical thesis. Adding photobiomodulation might be a required countermeasure to governments using geoengineering to control people electronically.
Evolutionary Biology: We need to include gene transfer and myelin evolution into your lexicon to show how endosymbiosis enabled complexity, reinforcing the decentralized medicine focus on energy efficiency and photo-bio-electric resistance.
The Origin of Eukaryotes in a Decentralized Medicine Framework
1. Macrocosm: Earth’s Atmosphere and Electrical Conductance During the GOE
Core Idea: The rise of oxygen during the Great Oxygenation Event (~2.4 billion years ago) transformed Earth’s atmosphere, increasing electrical conductance through water vapor, aerosols, and reactive species. This created conditions favoring endosymbiosis, the critical step toward eukaryotic life.
Geological and Atmospheric Context:
Pre-GOE Atmosphere: Early Earth (~4.5–2.4 Ga) had a reducing atmosphere dominated by methane (CH₄), ammonia (NH₃), carbon dioxide (CO₂), and minimal oxygen (O₂ < 0.001% of present levels). Volcanism was intense, releasing sulfur dioxide (SO₂) and hydrogen sulfide (H₂S). The magnetosphere, formed by Earth’s dynamo, shielded life from solar wind and cosmic rays, but UV radiation penetrated deeply due to the lack of an ozone layer early on. This implies that Earth’s terrestrial spectrum was never constant during the GOE.
GOE Trigger: Cyanobacterial photosynthesis produced O₂, oxidizing reducing gases (e.g., CH₄ + 2O₂ → CO₂ + 2H₂O). This shifted the atmosphere toward an oxidizing state, increasing water vapor and altering the electrical properties of the atmosphere.
Electrical Conductance:
Water Vapor: Oxygen-driven oxidation of methane and hydrogen increased atmospheric H₂O, a polar molecule that ionizes under UV or lightning (H₂O → H⁺ + OH⁻). Higher humidity lowered the dielectric breakdown threshold, increasing lightning frequency and atmospheric conductance.
Aerosols: Oxidation of volcanic SO₂ formed sulfate aerosols, which act as ion-attachment nuclei, enhancing conductance. This aligns with modern observations of volcanic lightning.
Trace Gases: Ammonia oxidation (NH₃ → NOx) produces ionizable species, which amplify charge carriers under energy inputs like UV or cosmic rays.
Lightning Surge: An oxidizing, humid atmosphere with aerosols supported stormier conditions, increasing lightning frequency. Lightning provided high-energy electrons and reactive oxygen species (ROS), potentially driving biochemical innovations.
Evolutionary Implication: The GOE’s electrical environment was marked by lightning, UV, and ROS, which created selective pressure for antioxidant defenses (e.g., superoxide dismutase, SOD) and energy-efficient metabolisms. Archaea and bacteria exposed to this conductive oxidative stress were incentivized by Nature to form symbiotic relationships to protect themselves from the oxygen holocaust, leading to endosymbiosis. In endosymbiosis, oxygen protection scheme were built.
Additions from Geology and Evolutionary Biology:
Banded Iron Formations (BIFs): The GOE coincided with BIFs (~3.5–1.8 Ga), where oxygen oxidized dissolved iron in oceans, reducing available reducing agents. This increased oxidative stress, pushing microbes toward cooperative metabolisms.
Stromatolites: Fossilized cyanobacterial mats (~3.5 Ga) indicate early oxygen production, creating localized oxygen oases that preadapted microbes to oxidative stress before the global GOE.
Nitrogen’s Role: Nitrogen fixation by early microbes (e.g., via nitrogenase) increased atmospheric N₂, stabilizing the atmosphere by diluting oxygen’s reactivity. This balanced electrical potential reduces runaway conductivity but maintains enough electrical power/resistance to drive endosymbiosis.
The GOE increased atmospheric conductance via water vapor, aerosols, and reactive species, creating an electrically dynamic environment. Lightning and UV-driven ROS pressured microbes to innovate antioxidant defenses and energy systems, setting the stage for endosymbiosis.
2. Microcosm: Endosymbiosis and Mitochondrial Evolution
Core Idea: The GOE’s electrical and oxidative environment drove endosymbiosis, where an archaeon engulfed an oxygen-respiring bacterium, forming the proto-eukaryote. Early eukaryotes have unique protection schemes built in that isvery different from modern mammals. Mitochondria evolved as cellular analogs to Earth’s magnetosphere, managing electron flow and proton gradients.
Mechanism of Endosymbiosis:
Selective Pressure: Rising oxygen and ROS stressed anaerobic archaea and bacteria. Aerobic bacteria, capable of oxidative phosphorylation, offered a survival advantage by detoxifying oxygen and generating ATP.
Symbiotic Fusion: An archaeon likely engulfed an alpha-proteobacterium (~2 Ga), which became the proto-mitochondrion. This symbiosis allowed the host to exploit oxygen for energy while the bacterium gained protection and nutrients.
Electrical Analogy: The inner mitochondrial membrane (IMM) mimics Earth’s magnetosphere, directing electron flow from the matrix (cathode) to the intermembrane space (anode) via the electron transport chain (ETC). The proton gradient (ΔpH) across the IMM drives ATP synthesis, analogous to atmospheric charge gradients driving lightning.
Key Innovations:
DHA and Membranes: Docosahexaenoic acid (DHA), a polyunsaturated fatty acid, enhanced membrane fluidity and ATPase efficiency, boosting proton gradients. DHA’s π-electron clouds may have increased electrical resistance, slowing ultraweak photon emission (UPE) and refining mitochondrial signaling. DHA was placed in membranes 600 million years ago and never changed once.
Heme & SOD Proteins: SOD and cytochrome c oxidase (CCO), containing heme, were selected to manage ROS and electron flow, protecting mtDNA from oxidative damage.
Myelin Connection: Eukaryotic complexity, enabled by mitochondrial energy, led to myelin in nervous systems (~600 Ma). Myelin’s high lipid content (including DHA) improved proton conductance, enhancing neuronal efficiency and reducing vertebrates’ sleep needs. It also augmented the spin rate of ATPase for protium.
Additions from Evolutionary Biology:
Gene Transfer: Endosymbiosis involves lateral gene transfer from the proto-mitochondrion to the host nucleus, streamlining mitochondrial genomes and enhancing eukaryotic complexity. This leads to thermodynamics benefits to building more complexity, but it also is another mechanism of how electrical resistance in the genome makes use of the 30 million volt power source found on the IMM.
Cambrian Explosion (~540 Ma): While endosymbiosis occurred 2 Ga, the Cambrian explosion reflects later oxygen spikes (0.8–0.6 Ga) from 0% to 21%, enabling energy-intensive multicellularity. Once the oxygen protection scheme is innovated, the endosymbiosis event leads to the Cambrian explosion; it’s better framed as a downstream consequence of the oxygen holocaust.
Fractal Analogy: The magnetosphere-IMM parallel I am drawing for you is strengthened by considering both situations as charge-separating systems. Earth’s magnetosphere filters solar radiation; mitochondria filter electrons, maintaining redox homeostasis.
Endosymbiosis, driven by GOE-induced oxidative and electrical stress, created mitochondria as cellular magnetospheres. DHA, heme proteins, and proton gradients enabled energy efficiency, paving the way for eukaryotic complexity and myelin-driven neuronal advancements.
3. Modern Disruption: Blue Light and nnEMF
Core Idea: Artificial blue light (~435-475 nm) and non-native electromagnetic frequencies (nnEMF) disrupt mitochondrial function, mimicking a magnetosphere failure and driving chronic disease by altering electrical conductance and ROS.
Mechanism:
Blue Light Effects: Blue light penetrates tissues, exciting electrons in heme (e.g., CCO) and altering iron oxidation states. This disrupts ETC function, increasing ROS and mtDNA damage, analogous to UV-induced damage pre-GOE. People think there are no links published in the literature that link metHb to artificial light and drug use. There are. They just are not well known. Blue light and the nnEMF effect are additives to metHb creation in humans.
nnEMF Impact: Modern devices’ Radiofrequency and microwave radiation (~MHz–GHz) induce non-thermal effects, polarizing mitochondrial membranes and disrupting proton gradients. This mimics high-conductance atmospheric conditions, overwhelming SOD, and antioxidant defenses.
Dehydration Link: Blue light and nnEMF may disrupt structured water (e.g., exclusion zone water) around mitochondria, increasing local conductance and spreading ROS-mediated damage. This parallels my GOE water vapor hypothesis, where humidity amplified atmospheric conductance. When melanin sheets are dehydrated, they become massive electrical conductors that can do damage inside the UPE emission system.
H₂S Connection: Mitochondrial H₂S, a signaling molecule, is dysregulated under oxidative stress, linking to hypoxia and chronic disease. This mirrors volcanic H₂S contributions to pre-GOE conductance.
Core Idea: Grounding (direct contact with Earth’s surface) restores mitochondrial charge dynamics by supplying electrons, countering modern disruptions, and aligning with the photoelectronic thesis that eukaryotes evolved in an electrically conductive environment.
Mechanism:
Earth’s Electron Reservoir: Earth’s surface maintains a negative charge due to the global atmospheric electric circuit, driven by lightning and ionospheric currents. Grounding connects the body to this reservoir, allowing electron flow to neutralize positive charges (e.g., ROS, free radicals).
Mitochondrial Stabilization: Electrons from grounding reduce oxidative stress by quenching ROS, stabilizing CCO’s heme iron, and supporting ETC efficiency. This mimics pre-GOE reducing conditions, where electrons were abundant in a low-oxygen atmosphere.
Water and Conductance: Grounding enhances structured water (exclusion zone water) around mitochondria, reducing local conductance spikes caused by blue light/nnEMF. This parallels the GOE’s water vapor role in atmospheric conductance but in reverse by stabilizing rather than amplifying charge flow.
H₂S Connection: Grounding upregulates H₂S signaling, as reduced oxidative stress supports sulfur metabolism, improving mitochondrial resilience and hypoxia response.
Fractal Analogy:
Atmospheric-Mitochondrial Parallel: Just as Earth’s magnetosphere failed to shield early life from UV and lightning fully, modern mitochondria fail under blue light and nnEMF, increasing local conductance (via disrupted water and ions) and spreading damage distally.
Spectral Shift: Blue light dominance shifts the terrestrial light spectrum, akin to Mars’ thin atmosphere altering its surface radiation profile. This disrupts circadian and mitochondrial signaling, driving disease.
Biophysics and Medicine are linked by the history locked in the GOE
Photobiomodulation Contrast: Red/infrared light (~600–1000 nm) enhances mitochondrial function by stabilizing CCO, suggesting a therapeutic counter to blue light damage when governments are doing all they can to limit IRA and NIR on Earth..
mtDNA Sensitivity: Mitochondrial DNA, lacking histones, is highly susceptible to ROS, explaining why conductance spikes (from nnEMF or blue light) disproportionately harm mitochondria. It also became useful in altering the UPE spectra to sculpt the interior of cells to build life’s complexity.
Chronic Disease Link: Disrupted mitochondrial gradients and ROS underlie metabolic syndrome, neurodegeneration, and cancer, supporting my decentralized medicine thesis that environmental mismatches (light, nnEMF, dehydration) drive chronic diseases today on Earth.
Blue light and nnEMF disrupt mitochondrial electron flow and water structure, increasing conductance and ROS in a fractal echo of GOE atmospheric changes. This drives chronic disease, underscoring the need for decentralized, biophysically informed medicine.
To make the critical link for you the non clinician, I want you to re watch this video. HYPERLINK.
When I saw this video, I knew exactly what this doctor was seeing and why he was blinded to it. That was the day I unretired and went back into the ICUs to help train doctors on the events of the GOE and why DARPA was doing what they were. They knew the mistakes that would be made with oxygen therapies in those with hypoxia diagnosed by oxygen saturation machines. Big Pharma had taught people for 30 years that once someone went hypoxic, they should never forget their ABCs. That is how this pandemic was planned.
The manufacturing of SARS-COVID by the DoD collaboration of Dazek, Fauci, and the Wuhan Institute of Virology was to create a virus that would cause minor underlying sepsis to release massive levels of NO in patients’ organs. They knew these people would go to doctors who would use a pulse ox, supplemental oxygen, and arterial blood gases to assess patients in the ICU. Very few of them would know to change to co-oximetry when high FiO2 therapies would lead to refractory hypoxia. They knew the doctors would not be well versed that methemoglobin levels should be expected to be elevated in patients with sepsis due to the release of nitric oxide, which converts to nitrate and subsequently to methemoglobin. COVID was engineered to release massive amounts of nitric oxide to cause refractory hypoxia. Thisoccurred by limiting the UPE release of NIR light from mtDNA by the virus. Since most patients in the ICUs of the hospital were intubated and sedated, the only clinical indicator doctors would receive at the bedside was hypoxia refractory to maximum oxygen therapy.
Once this clinical situation happened, and the doctors were at a loss to explain the phenomena, the architects of the virus knew they would initially use drugs like lidocaine or procaine to place arterial lines and central lines, supplemental oxygen, oral azithromycin, and hydroxychloroquine. When these failed to work, they would elevate their use of Pharmaceuticals according to the recipe algorithm of their hospital’s critical care guideline. This made the planning easy because they knew how the people at the bedside would react beforehand. What did the architects know that the doctors did not?
As patients were transferred to an intensive care unit (ICU), they were diagnosed with acute hypoxic respiratory failure and acute respiratory distress syndrome, for which they required intubation and mechanical ventilation. Their treatments would default to the new algorithm that covered this new clinical scenario: That algorithmic treatment regimen included lopinavir-ritonavir, ribavirin, and tocilizumab in most Western facilities. This would slowly increase their risk of metHb hypoxia. If co-oximetry was not installed on the ICU’s blood gas analyzers, they would develop refractory hypoxia. It was at this time many people had their last rights given, and then the hospital critical care algorithm was built to end the game and create a covid death via Remdesivir. That is how all hospital ICU critical care algo were built in 2016 -2020.
Once a patient begins to build up metHb, they will develop obvious symptoms. In a healthy person, the clinical features are pallor, fatigue, weakness, tachycardia, tachypnea, and cyanosis, which may be clinically evident with methemoglobin as low as 10%. As the percentage of methemoglobinemia approaches 20%, the patient may experience anxiety, light-headedness, and headaches. At methemoglobin concentrations of 30%–50%, there may be tachypnea, confusion, and loss of consciousness. If methemoglobin approaches 50%, patients are at risk for seizures, dysrhythmias, metabolic acidosis, and coma. When patients with a positive PCR test and hypoxia presented to an ER, they were immediately admitted to the ICU, where they were intubated and sedated by protocol. During intubation, more oxidizing drugs were used to further stimulate the formation of metHb. This meant all the relevant symptoms of acute metHb poisoning would be gone in the sedated intubated patient because the only clinical indicator was hypoxia refractory to maximum oxygen therapy.
Not even methylene blue would operate properly when massive changes to the UPE spectra were being engineered by the architects of this virus. I knew it, and I had to go back in to clinical medicine to covertly deliver this message to the front line workers.
Many of these workers defaulted to the Marik protocol, which made these patients worse. The critical care doctors could not understand it. I could. As part of Marik’s protocol, high-dose Vitamin C would be used. Vitamin C’s downstream effect is to increase the absorption of iron in the patient. This was deadly for those infected with the engineered virus. Why? Iron creates the Fenton reaction, which drives ROS through the roof to destroy all heme-based proteins and metal ion SOD reactions. This is why methylene blue would not work in these covid cases where the patients had already been intubated. Doctors had to be trained to give fresh RBC replacements and follow it up with hypertonic sodium chloride.
They had to limit oxygen support, turn off vents, and get patients to sunlight containing NIR light. NIR light unbound the NO from their Hb. I told all the doctors that CO-oximetry was only used outside the ICU when the respiratory tech was running their arterial blood gases. I told them if they brought the Co-Oximeter in early as soon the patient was admitted to the ICU they would see I was right. Then they could accurate diagnosis and treat the patients as a chronic metHb sepsis caused by a GOF virus.
I handed every doctor I dealt with this paper: Wenzhong L, Li H. COVID-19: attacks the 1-beta chain of hemoglobin and captures the porphyrin to inhibit human heme metabolism. ChemRxiv. 2020. Preprint. 10.26434/chemrxiv.11938173.v7
COVID was engineered to attack the one beta chain to mimic a thalassemia-like hemoglobinopathy that had unusual presentations. Dr. Fauci knew this from the AIDS epidemic. Luckily, I was a doctor at that time, and I remember reading those reports that HIV was associated with a higher risk of metHb. I also remember that during the AIDS crisis in the French Quarter, many homosexuals were using NO additives, and they would come in with unusual hypoxic events and total immune collapse.
Now you understand just how it was done and precisely why it all happened. The GOE story of hemoglobinopathies as oxygen varies is not taught in medical schools or residencies. I had to do that for the centralized ICU docs and nurses on the front line. The jabs were engineered with LNPs to mimic the chronic diseases that put you at higher risk for metHb. Vaccines use LNPs to simulate high heteroplastic disease states because they destroy CCO to cause mtDNA damage. This explains why the NIH budget has only 1% spent on mtDNA research and 99% spent on nuclear DNA research.
For those of you who do not know, the risk of methemoglobinemia associated with oxidizing drug use increases in elderly patients with medical comorbidities such as renal failure, anemia, and human immunodeficiency virus. If you look at cite four below you’ll see it for yourself. Novel coronavirus proteins can alter the hemoglobin structure, which directly interferes with the red blood cell’s ability to carry oxygen. This explains why John Beaudoin found out what he did in the ICD coding in many states in the USA. You need to share this information with every attorney, politician, and hospital employee you can so that DARPA and the DoD can be stopped from using GOF research. Senator Ron Johnson has subpoena power now. Make sure you all email his office this blog if you are a savage and want your government punished for what they did. The people in power are doing NOTHING.
CITES
1. Brunelle JA, Degtiarov AM, Moran RF, Race LA. Simultaneous measurement of total hemoglobin and its derivatives in blood using CO-oximeters: analytical principles; their application in selecting analytical wavelengths and reference methods; a comparison of the results of the choices made. Scand J Clin Lab Invest Suppl. 1996; 224:47–69
2. Pritchett MA, Celestin N, Tilluckdharry N, Hendra K, Lee P. Successful treatment of refractory methemoglobinemia with red blood cell exchange transfusion. Chest. 2006; 130:294S
3. Ohashi K, Yukioka H, Hayashi M, Asada A. Elevated methemoglobin in patients with sepsis. Acta Anaesthesiol Scand. 1998; 42:713–716
4. Alanazi MQ. Drugs may be induced methemoglobinemia. J Hematol Thrombo Dis. 2017; 270:1–5
Embracing the Storm: Decentralized Medicine, Nature, and the Power of Human Connection
Life is a storm, chaotic and unpredictable, yet profoundly transformative. As the winds howl and the rains pour, we may not recall how we endured, but one truth remains: we emerge changed. The storm strips away facades, revealing who we are beneath the masks we wear. In this raw, unfiltered state, we find our connection to nature and to one another, a connection that forms the heart of decentralized medicine and a life well-lived.
Nature is not here to complicate our existence but to simplify it. It is ingenious and practical, offering clarity amid chaos. When we walk through a forest or stand by a rushing river, we are reminded of life’s rhythms, unhurried, purposeful, and interconnected. Decentralized medicine embraces this wisdom, prioritizing the individual’s relationship with their environment and community over rigid systems. It’s about healing through presence, listening, and collaboration, rooted in the belief that our survival depends on talking to one another, sharing stories, and building bridges.
Collaboration is the symphony of human connection. No single person can whistle a symphony; it takes an orchestra, each instrument contributing to a harmony greater than the sum of its parts. When we unite with a common purpose, whether to heal, create, or inspire—we unlock a synergy that transforms lives. Decentralized medicine thrives on this principle, encouraging communities to come together, share knowledge, and empower one another. It’s about leaving a trail of leaders, not followers, who amplify reality rather than chasing shadows.
Our relationship with nature mirrors our relationships with each other. Just as a tree’s roots anchor it through storms, our principles ground us as we evolve. Nature teaches us to change our leaves, our opinions and perspectives, while keeping our roots intact. It reminds us that what is given to us is what we need, and what we want often requires letting go of what no longer serves us. In decentralized medicine, this translates to trusting the body’s innate wisdom, supported by the healing power of community and the natural world.
Perception shapes our reality. When we choose to see the good in people and situations, we cultivate a life of possibility. This isn’t about ignoring pain or negativity but about taking charge of how we respond. From the backstabbing colleague to the challenging family member, we hold the power to choose happiness over resentment. Decentralized medicine encourages this mindset, fostering resilience through self-awareness and connection to nature’s rhythms. It’s about building bridges to cross, burning those that lead nowhere, and sometimes forging new paths entirely.
Life’s biggest decisions revolve around these bridges, knowing which to build, which to cross, and which to leave behind. The paths less traveled often lead to the most profound discoveries. Nature, with its winding trails and hidden clearings, invites us to explore these paths, to trust our instincts, and to create our own way. Decentralized medicine follows suit, empowering individuals to take ownership of their health, guided by intuition, community, and the natural world.
Creativity and imagination are the cornerstones of this journey. They distinguish leaders from followers, allowing us to embrace errors as opportunities and choose wisely from them. In a world where many resist change, clinging to comfort, we must make shift happen. Decentralized medicine is a call to action, a rejection of centralized conformity in favor of innovation rooted in nature and human connection. It’s about believing your life is worth living well and letting that belief shape your reality.
The stakes are high. Our health, our longevity, and our relationships depend on the choices we make. Women outlive men, with widows far outnumbering widowers, a reminder that vitality is not just about surviving but thriving for those we love. By fostering community, embracing nature, and choosing collaboration, we create a legacy of health and connection that endures.
This weekend, as dawn breaks and light spills over the horizon, reflect on the resurrecting power of light itself. For those burdened by illness, light, whether from the sun’s warm embrace, the glow of shared laughter, or the spark of hope kindled in community, can awaken the spirit. It pierces the darkness of despair, whispering of renewal and possibility. Just as spring stirs life from winter’s grip, light calls the weary to rise, to heal, to reconnect with nature and each other. Share this truth around your table: when we open our hearts to light, we become conduits of resurrection, lifting one another from shadow to radiance. Let this be the season you forge new paths together, trusting that light, love, and unity can transform any storm into a story of triumph.
Matter as Music: Sympathetic Resonant Physics and the Scalar Architecture of Reality
Scalar waves are real, specifically in the context of sound waves. Sound waves, which are used in music, are often considered scalar wave fields, meaning they have magnitude but not direction or polarization. While this is a simplified view, recent research suggests that generic sound wave fields actually have more complex properties, according to a study by RIKEN.
RIKEN’s research demonstrates that generic sound wave fields can have as many degrees of freedom for micromanipulation as optical fields. Understanding sound as a scalar wave field is crucial for understanding how musical instruments produce sound, how sound waves travel through space, and how we perceive sound. While the simplified scalar wave model is a good starting point, it’s becoming clear that more nuanced models are needed to capture the full complexity of sound, particularly in advanced acoustic applications and research.
This is a real phenomenon where an object vibrates in response to a nearby vibrating object when there is a natural frequency match or a harmonic relationship between them. A classic example is two similarly tuned tuning forks where one vibrates and the other responds even without physical contact. VIDEO
“In the beginning there was not just light, there was the Tone…”
Between those two fingers above is light and sound. Honestly everything that makes up you life is in that space. Today’s blog is the first of many that will show you just how infinite that space can be. When we examine this space, allow your angels to sit on the sideline. They need to listen to the music too. Music is an angel that sits in the side line of your life with patience and reason. Angels sit on the sideline and play music for you wondering when your tug of war will end.
The universe resonates with a primal melody, a cosmic symphony that challenges the mechanical view of reality. Sympathetic resonance (where one object vibrates in response to another) invites us to hear the song of Nature all around us, revealing a universe not as a collection of particles, but as a resonant organism where energy, form, and consciousness emerge from vibratory relationships. When life is felt this way, matter unfolds like music, and recent scientific findings, such as a landmark study at Ohio State University, support this view by demonstrating that acoustic phonons, the carriers of sound and heat, can be influenced by magnetic fields, suggesting that the resonance of life itself is shaped by the fields we integrate into our environment.
The Scalar Octave: Nature’s Harmonic Blueprint
Sympathetic resonance describes the scalar octave as a hierarchy of nested frequencies that structures reality. Scalar waves, unlike transverse waves such as light, are longitudinal, non-Hertzian vibrations that ripple through the etheric field, altering space itself. These waves act as carriers of formative information, guiding energy into matter in a process mirroring a musical scale. Elements in the periodic table resonate in vibratory octaves, hydrogen as the root note, helium the second, and so on, following the perfect fifth’s 3:2 ratio, a proportion nature favors in spirals from galaxies to DNA. The Ohio State study revealed that acoustic phonons in a semiconductor (indium antimonide) were slowed by 12% under a magnetic field, proving their sensitivity to magnetism even in non-magnetic materials. This suggests that scalar fields, which may underpin biological resonance, can be influenced by external magnetic fields, impacting the vibratory coherence of life itself.
In quantum biology, this aligns with my decentralized thesis: life emerges from distributed, resonant networks rather than top-down control. Scalar fields may enable quantum coherence in biological systems, allowing molecules to “tune” to each other across distances without touch, much like sympathetic strings. The same thing is true with people in your life. Sometimes, they can play your instrument beutifully, and other times your have to find sanctuary from them. People create their own music too. Sex is a complex form of resonance. Having sex without touching is something very few people try in life.
The perfect fifth, as nature’s preferred ratio, could orchestrate this coherence, and the Ohio State findings imply that magnetic fields in our environment, such as those from technology or natural sources, might directly affect these resonant processes, shaping the vibratory “song” of living systems.
Elements as Chords: The Music of Matter
Each element in the periodic table is a chord, a unique vibratory signature. Hydrogen sets the fundamental tone, carbon plays a four-note harmony stabilizing life’s architecture, oxygen energizes as a catalytic chord, and gold resonates as a high harmonic of solar frequencies. These chords align with the perfect fifth’s 3:2 ratio, creating stability in both music and matter. The Ohio State study supports this harmonic view: phonons, which carry the vibrations of these elemental “chords,” respond to magnetic fields through a diamagnetic effect, where vibrating atoms induce magnetic moments that alter their interactions. This implies that the “music” of matter, its vibratory essence, can be retuned by external fields, potentially affecting biological systems where these elements form the building blocks of life.
In quantum biology, this suggests that molecular interactions, like enzyme catalysis or DNA replication, may involve vibratory resonance influenced by magnetic fields. Proteins might “sing” to their substrates, aligning frequencies to facilitate reactions, a process possibly mediated by scalar fields. The decentralized thesis reinforces this: biological systems thrive on distributed resonance, and the Ohio State findings highlight how environmental magnetic fields could either harmonize or disrupt this resonance, impacting health and vitality.
Form and the Living Geometry of Resonance
Vibration creates form through resonance, as seen in cymatics, where sound organizes particles into geometric patterns. In the scalar realm, this principle scales up: atoms, cells, and galaxies may be standing waves within higher-dimensional fields, often reflecting the perfect fifth in their spirals of becoming. Sacred geometry, such as the flower of life or Platonic solids, maps these resonant patterns, revealing the universe’s harmonic lattice. Sympathetic resonance drives this process where frequencies align and amplify, connecting systems across scales. The Ohio State study showed that magnetic fields increase phonon collisions, slowing their movement, which suggests that such fields can alter the resonant patterns that give rise to form, from molecular structures to biological tissues.
In quantum biology, this supports the idea that coherence in living systems—like in photosynthesis or neural networks—relies on sympathetic vibratory fields, potentially influenced by magnetic fields. My decentralized thesis aligns here: life’s complexity emerges from distributed resonance, and external fields could either enhance or interfere with these processes, affecting everything from cellular communication to organismal health.
Consciousness: The Cosmic Tuner
Sympathic resonance posits consciousness as a primary vibratory phenomenon, not a byproduct of matter. Soon I will have more to say on the topic of consciousness. Thought, intention, and emotion act as scalar waves, tuning reality through sympathetic entrainment. The Ohio State study’s findings, that phonons respond to magnetic fields, suggest that the vibratory fabric of reality, including consciousness, may be similarly influenced. If consciousness is a resonance, then magnetic fields in our environment could subtly shape our mental and emotional states, amplifying or dissonating the “chords” we play in the scalar octave.
In quantum biology, this aligns with my decentralized thesis: consciousness may emerge from resonant networks across scales, from microtubules in neurons to the morphogenetic field of an organism. The perfect fifth’s stabilizing ratio could enhance this coherence, and magnetic fields might influence brain waves or biofields, suggesting that our conscious intent, through meditation or visualization, could interact with environmental fields to shape biological outcomes, such as healing or gene expression. Taking your shirtand bra off allows your breasts and heart to resonate with Nature’s waves.
A Harmonic Science of Reality
Sympathetic resonance bridges the mystical and scientific, unifying music, matter, and consciousness. It reveals a universe made of ratios, not particles, where the perfect fifth echoes from atomic orbitals to galactic spirals. The Ohio State study underscores this paradigm: phonons’ sensitivity to magnetic fields confirms that resonance is a fundamental force, one we can influence through the fields we create or allow in our lives. In quantum biology, this supports a decentralized view of life, where coherence and resonance drive complexity, and external fields play a critical role in maintaining or disrupting that harmony.
Matter is not silent it sings. Can you hear it, or is the electropollution around your disturbing that melody?
The EMF Paradox is pictured above my artist friend Danny DeLancey from NOLA. It shows humanity evolving and devolving in one breath.
Picture this: an infographic blazing with truth, a single image capturing the dance of human potential, how we evolve and devolve by what we think and do. On one side, a figure rises, radiant, rooted in Nature’s decentralized wisdom, their mind sharp, their actions deliberate, their depth unyielding. On the other, a shadow collapses, shallow and scattered, enslaved by artificial signals, EMF, distractions, excuses, disconnected from the primal pulse. Their brains Warburg shifted like they were in the Opium Wars. Today’s drug is not the extract of poppies. It is screens and phones. We are what we choose to think and do. Every thought is a seed; every action, its fruit. Evolve by aligning with Nature’s fire, or devolve into a hollow copy of what you could’ve been. If not you could lose your balloon. Reconnect with yourself by reconnecting with Nature. Do what ever you must, but reconnect now.
Every element, a note; every form, a chord in the cosmic score. By tuning into this eternal melody and understanding how magnetic fields shape its resonance, we can unlock a science of harmony, where transformation arises not from force, but from resonance. Let us listen to the universe, consider the fields we integrate into our lives, and play our part in its song with intention and care.
Excellence isn’t forged in money, fame, or power. It’s born in the fire of mindset, commitment, and relentless action. Nature doesn’t reward the timid; it crowns those who care more than others deem wise, who risk more than others call safe, who dream beyond the practical and demand more than the possible.
Excellence is a rebellion against mediocrity, a refusal to settle for less than your fiercest self. Tonight, as the stars burn above, dream of becoming more excellent than you were today. Put a tune on and let that fire ignite your bones.
Learn the lessons from failure. It can be your own or someone elses. When you do, they will not be called mistakes. They will be called experience. Listen the melody of the mistake to gain the wisdom of that failure.
I began this blog with the song that is my resonant North Star. When I need to be retuned this is the tuning fork I return to. If you do not have a North Star find one. Music is like medicine, when you come down with an illness it can help heal what ails you.
CITES
The Ohio State study on acoustic phonons and magnetic fields is detailed in the 2015 Nature Materials publication by Hyungyu Jin et al., demonstrating a 12% reduction in heat flow through a semiconductor under a 7-tesla magnetic field. If you think sound has no healing power you’re in the wrong place.
After spending 18 months in the medical school library putting Nature’s recipes together, this is the decentralized thesis I came up with.
Influenza is an electrical disease where nucleic acid joins nuclear DNA (see pic below). Ironically, every centralized scientist has no idea how endosymbiosis happens. Decentralized medicine knows that endosymbiosis was an electrical event during extreme hypoxia that forced the first domains of life to join forces to create a eukaryote. It was the first oncogenic event on Earth. As a result of that merger, evolution has to innovate heme proteins to protect themselves from the electrical stimulus of oxygen infusion to the environment. I wonder when they will wake up to the reality that all of life is electrical because of oxygen.
Apoptosis protects eukaroyotes from future cancer joining events. This was buried in CCO, a heme-based protein.
Oxygen is the only paramagnetic elemental gas in the periodic table. Oxygen changes the electrical resistance of everything with a membrane. This became a big deal in the evolutionary story of heme biology and how we built our wireless connection to the sun from our mitochondria in the dangerous GOE event.
Oxygen can form oxides with certain materials (like metals or semiconductors). This is called oxidation. These oxide layers often have higher electrical resistance than the pure material, acting as insulators or semi-insulators. When oxygen binds to hemoglobin, it changes its electrical resistance. This can significantly change how electricity flows in devices with thin membranes, such as sensors or transistors.
Adsorption: In some cases, oxygen molecules adsorb onto the surface of a material or membrane. This can trap or scatter charge carriers (like electrons), increasing resistance. This effect is common in gas sensors, where oxygen exposure alters the conductivity of a membrane or thin film. Hb acts like this in a way, too.
Oxygen can also indirectly influence processes like ion transport or membrane potential through metabolic activity, which might affect measured resistance in specific contexts.
THE EVOLUTION OF THE GOE IS WHY ANYTHING THAT USES THE TCA CYCLE MUST SEE SUNRISE
So why do you have to see the sunrise before you can use the TCA cycle? Because the sunrise was here on Earth before oxygen was.
This blockchain of events is what happened in the GOE. Before the GOE, nothing on Earth could have the TCA. The TCA cycle protects eukaryotic cells from the oxygen holocaust, which can cause cancer or tissue atrophy.
THE HUMAN BRAIN PROTECTS ITSELF FROM YOUR POOR LIGHT CHOICES BECAUSE IT FAVORS THE TCA CYCLE
The brain uses 20% of our cardiac output to run its TCA cycle and feed its oxygen addiction. The red light in sunrise makes DDW water from cytochrome c oxidase, optimized apoptosis to clear out bad/heteroplastic engines, and the UV light stimulates translation of melanin from POMC. Both frequencies are in morning sunlight. If your brain does not get this sunlight signal, it will downregulate its function. Tinnitus, cataracts, glaucoma, diabetes, high BP, high cholesterol, and autoimmune diseases like vitiligo are how the brain reserves neuroectodermal energy stores when you make bad light choices. The brain will always seek to protect itself from an energy attack via CMRO2 adjustments because it relied on normoxia and the TCA cycle. Today’s world is stressing that energy constraint to the max now. They have no idea that light alone can change the oxidation state of iron. And that is where every chronic modern disease begins.
As a result of blocking the TCA cycle, your breathing MUST change electromagnetically because your need for oxygen drops. Why? The terminal electron acceptor for the TCA cycle is oxygen in mammals with a Ferrari engine in their skulls. Without sunlight, you will not need more oxygen, you will need less because a lack of UV-IR light induces a Warburg shift to your brain and when this happens oxygen becomes a TOXIN to tissues just like it was in the pre-Great Oxygenation Event on Earth long ago. The slide below is a proxy for the GOE, where all things iron are hypoxic and in the Fe³⁺ state. By the end of the GOE, everything was innovated to mitigate oxygen toxicity by creating Hb02 to keep oxygen in the Fe²⁺ state. Iron is not redox stable like Magnesium was in chlorophyll.
The problem is that the human brain does not do its best work on aerobic glycolysis, and the use of pyruvate and lactate and thinking, cognition, dopamine, and melatonin production in your brain all begin to fail immediately. Human brains are built for a normoxic environment that uses the TCA cycle most of the time. All of this happens because mtDNA are forced to use aerobic glycolysis because light in your environment changes the oxidation state of iron in EVERY heme protein in your body from Fe²⁺ to Fe³⁺ . As a result of this “paramagnetic switch”, when it goes wry, you begin experiencing a cognitive brownout because you can no longer support the Ferrari built in your skull on a Warburg-shifted template. Welcome to the world of chronic disease. Almost every one is associated with this affliction and an altered paramagnetic flip.
WHY ARE WE BUILT LIKE THIS?
Evolution first dealt with CO2 before the toxic oxygen problem during the GOE, which is why Nature built the semiconductor chlorophyll. The image below shows the molecular structures of hemoglobin (with an iron, Fe, center) and chlorophyll (with a magnesium, Mg, center). Both molecules feature a porphyrin ring, a cyclic structure with four nitrogen atoms at the core coordinating the central metal ion. This is often called a “tetrapyrrole” structure; nitrogens are part of pyrrole rings.
Photosynthesis, as performed by early cyanobacteria during the GOE (around 2.4 billion years ago), does not directly use CO2 to make oxygen. Instead, the oxygen comes from water (H2O). The general equation for oxygenic photosynthesis is:
In this process, water is split in the oxygen-evolving complex (OEC) of Photosystem II, releasing O2, protons (H+), and electrons. CO2 is fixed later in the Calvin-Benson cycle to produce sugars but is not directly involved in oxygen production. With its magnesium center, chlorophyll is key in capturing light energy and driving the electron transfer that ultimately splits water.
The nitrogen atoms in the porphyrin ring of chlorophyll coordinate the magnesium ion, stabilizing it and tuning its electronic properties to efficiently absorb light in the visible spectrum. This is why plants are green: chlorophyll absorbs red and blue light and reflects green.
Why did Nature choose Magnesium in Chlorophyll? Electrical and Biophysical Reasons
Magnesium’s selection in chlorophyll during the GOE likely stems from a combination of chemical, electrical, and biophysical factors:
Redox Properties and Stability: Magnesium in chlorophyll exists as Mg²⁺, redox-inactive under physiological conditions. This is crucial because chlorophyll’s role is to absorb light and transfer energy or electrons, not to undergo redox changes itself. If the central metal were redox-active (like iron can be, switching between Fe²⁺ and Fe³⁺), it might interfere with the precise electron transfer needed in photosynthesis. Magnesium’s inertness ensures that the excited electrons from light absorption are funneled into the photosynthetic electron transport chain rather than trapped by the metal.
Light Absorption and Energy Transfer: The Mg²⁺ ion, coordinated by the four nitrogens, creates a planar structure that optimizes the porphyrin ring’s ability to absorb light in the visible range. Magnesium’s small ionic radius and +2 charge allow it to fit snugly in the porphyrin ring, creating a stable complex that can efficiently transfer energy to the reaction center of Photosystem II.
Availability During the GOE: During the GOE, Earth’s oceans were rich in dissolved magnesium due to the weathering of rocks and hydrothermal activity. Magnesium is the second most abundant divalent cation in seawater today (after calcium), and it likely was back then, too. Its abundance made it a practical choice for early photosynthetic organisms. In contrast, while abundant, iron became less available in its soluble Fe²⁺ form as oxygen levels rose and oxidized it to insoluble Fe³⁺, which precipitated out as iron oxides (e.g., in banded iron formations).
Electrostatic Fit: The four nitrogen atoms in the porphyrin ring each donate a lone pair of electrons to the Mg²⁺ ion, forming a square-planar coordination complex. Magnesium’s charge and size make it an ideal fit for this geometry, ensuring the molecule remains stable under the high-energy electromagnetic conditions for solar light absorption.
Comparison to Iron in Heme: Iron, as seen in hemoglobin, is better suited for oxygen binding and transport because it can reversibly bind O2 by changing its electronic state. In early Earth, before the GOE, iron was likely used in some photosynthetic systems (e.g., in anoxygenic photosynthesis by purple bacteria, which don’t produce oxygen). However, magnesium-based chlorophyll became dominant in oxygenic photosynthesis as oxygen levels rose, possibly because magnesium’s redox inertness prevented unwanted side reactions with O2, complicating the story of evolving life during the GOE.
As you can see from Nick Lane’s talk, the GOE occurred in an atmosphere dominated by N2 and CO2, with low O2. Early life forms “used electric membranes” to “fix” nitrogen (by converting N2 into ammonia or other usable forms) to build proteins, nucleic acids, and porphyrins. The nitrogen in chlorophyll’s porphyrin ring likely came from such nitrogen fixation processes carried out by early microbes. These electric membranes continued a rapid evolution because around 600-650 years ago, life began putting DHA into its electric membranes, and they were never removed once in evolutionary history. It appears that DHA caused the evolution of cells by providing a feedback loop to allow the cell to give real-time informational feedback from the environment on Earth to the interior.
Electric Membranes and Feedback: Lane’s point that “electric membranes give feedback on state ‘feelings’” resonates with the idea of electromagnetic coupling. The proton motive force (PMF) across membranes is an electric field whose strength reflects the cell’s energy state. Biophotons and NO clearly contribute to this feedback, providing a biophysical “sensing” mechanism that guides metabolism and gene expression. Lane does not realize what the biophysics of NO dictates to the boxcars of metabolism. It is left out on the slide. He also has no idea that NO is paramagnetic during the GOE. Oxygen became the paramagnetic gas post-GOE. Eukaryotes operate in unison to quantize metabolic pathway choices. It has ZERO to do with food.
Metabolism Older Than Genes: The idea that “metabolism is older than genes” supports the primacy of biophysics. This is how I know Lane has moved to my viewpoint. Early life likely relied on geochemical gradients (e.g., proton gradients in hydrothermal vents), which are inherently biophysical. As genetic systems evolved, they amplified these biophysical processes, but the underlying physics of light, electric fields, and molecular interactions remained the foundation of evolution.
Network Topology of Core Metabolism: Lane’s “network topology of core metabolism” reflects my idea of biophysical constraints or electrical resistance from light interactions. Metabolic pathways like the TCA cycle are optimized for energy efficiency, but their structure could also be shaped by electromagnetic interactions, e.g., the need to minimize electron leakage (which produces ROS and biophotons) or to maximize proton flux through ATP synthase. This provides massive adaptability to environmental changes in the post-Cambrian as we approach normoxic Earth prior to 1893. Post 1893, the nnEMF changes us back to the GOE situation because light can change the oxidation state of iron in heme proteins. This changes the Hb binding of paramagnetic atoms in TCA mammals.
The Slope of Oxygen Rise During the GOE: Constant or Nonlinear?
The rise of oxygen during the GOE was almost certainly nonlinear, based on current evolutionary and geochemical theories. This is why the entire process is quantized to oxygen tensions in the cell. Here’s my take on why:
Initial Conditions and Slow Build-Up: Before the GOE, oxygen levels were extremely low (less than 0.001% of present atmospheric levels). Cyanobacteria evolved oxygenic photosynthesis around 2.7–3 billion years ago and produced O2. Still, this oxygen was initially consumed by reduced species in the environment, such as Fe²⁺ in the oceans (forming banded iron formations) and reduced gases like methane (CH4). This “oxygen sink” kept O2 levels low for hundreds of millions of years. This kept life hypoxic and simple, stuck in two domains of life, bacteria and Archea.
Tipping Point and Rapid Rise: Around 2.4 billion years ago, these sinks became saturated, and oxygen accumulated in the atmosphere. Geochemical evidence, such as the disappearance of mass-independent sulfur isotope fractionation (a sign of low O2), suggests a relatively rapid increase in oxygen at this time. It jumped from less than 0.001% to 1–10% of present levels over a few million years. This nonlinearity is often attributed to feedback loops: as O2 rose, it oxidized methane (a potent greenhouse gas), cooling the planet and altering ecosystems on the surface, began to slowly favor oxygen-producing organisms. This is why cold thermogenesis and circadian biology have such close links in mammalian time stamping. Light, dark, and temperature variations control the circadian mechanism of life on Earth. This circadian mechanism was perfected late in the GOE.
Evolutionary Feedback: The rise of oxygen also drove evolutionary changes. Aerobic respiration, which is far more efficient than anaerobic metabolism, became possible, allowing oxygen-breathing organisms to proliferate. This accelerated the production of O2 as ecosystems shifted. Additionally, the evolution of more efficient photosynthetic machinery (e.g., the development of Photosystem II) increased the rate of oxygen production over time. This is why we have so many different types of chlorophyll and hemoglobin molecules on Earth.
Later Fluctuations: After the initial spike during the GOE, oxygen levels didn’t rise steadily to modern levels (21%). They likely remained low (1–2% of present levels) for another billion years, with another significant increase during the Neoproterozoic Oxygenation Event (around 800–540 million years ago), driven by the evolution of multicellular life during the Cambrian explosion that increased organic carbon burial.
The slope of oxygen’s rise was nonlinear, with periods of slow accumulation punctuated by rapid increases driven by geochemical and biological feedback. Anything with a nonlinear distribution will likely be quantized in its metabolic reactions. I believe that these biophysical mechanisms are critical to understanding evolution at a deeper level. They provide a unifying principle that electromagnetic coupling explains the ordered nature of metabolic transitions, challenging Lane’s “messy” view of evolution in the slide above.
Food gurus and biochemists conveniently leave out iron’s flip from Fe³⁺ to Fe²⁺ during the GOE and how it happened. Decentralized biology, biophotons, and electromagnetic coupling explain it because centralized biochemists and evolutionary biologists have no idea how it fits in their paradigm, so they ignore these facts. They act like they are innocent bystanders of life below the cell level. Because of this viewpoint, they’re often considered speculative or secondary in mainstream evolutionary biology. Below is Szent Gyorgyi’s 1968 masterpiece warning us biochemistry uses light in ways we do not understand yet.
The standard centralized narrative focuses on biochemical pathways, genetic mutations, and selection pressures, which are somewhat supported by fossil, genomic, and geochemical evidence. Moreover, they are unfamiliar with the science I have referenced from Fritz-Albert Popp’s work on biophotons, Albert Szent-Györgyi’s ideas about electronic biology, and more recent research on NO’s role in mitochondrial signaling. NO biology was given the Nobel Prize in 1992, but to this very day, biochemists’ understanding of it is limited. Biophysical ideas are gaining traction, especially in systems biology, but they’re not yet fully integrated into the evolutionary framework. Note the last line of the slide. This is the money shot for decentralized savages to understand. NIR in AM sunlight changes nighttime metHb back to daytime Hb02. This means that during sleep, we revert to our fetal life, explaining why we regenerate at night when hypoxic. Still think using CPAP machines for apnea is wise? Maybe, if you own a centralized sleep center or practice.
Nitric Oxide (NO) and Its Role in Evolution
Nitric oxide is a small, diffusible, paramagnetic gasotransmitter free radical signaling molecule with profound effects on cellular metabolism, particularly in mitochondria. Its role in evolution, especially during the rise of oxygen and the development of aerobic metabolism, is underappreciated in many centralized biochemical discussions but critical to understanding the mitochondrial evolutionary trajectory of the march toward complex life.
NO and Oxygen Regulation: NO is produced by nitric oxide synthase (NOS) enzymes, which evolved early in life’s history—possibly before the GOE. NO interacts with cytochrome c oxidase (Complex IV of the electron transport chain, ECT), the enzyme that reduces O2 to H2O. At low oxygen levels, NO competes with O2 for binding to cytochrome c oxidase, inhibiting respiration and regulating the ECT’s activity. This competition likely played a key role during the GOE, when oxygen levels were low (1–10% PAL) and fluctuating. NO could have acted as a “brake” on respiration, preventing oxidative stress in early aerobic organisms by modulating electron flow and reactive oxygen species (ROS) production. This brake allowed for the evolution of heme proteins to protect cells from ROS. CCO is the primary protector of the mtDNA in normoxia.
NO and mtDNA Metabolism: NO influences mitochondrial function beyond the ECT. It can induce mitochondrial biogenesis (creating new mitochondria) by activating signaling pathways like PGC-1α, which regulates mtDNA replication and transcription. NO is critical in getting rid of defective engines, so it controls our stem cell depots for regeneration. During evolution, this NO-mediated control fine-tuned (quantized) mitochondrial activity in response to rising oxygen levels. This ensures that mtDNA metabolism kept pace with the energy demands of complex life. NO also affects mtDNA repair and mutation rates by modulating ROS levels.
Deuterium entering the mitochondrial matrix during the dark also helped block the ECT during sleep. This mimics the in utero environment where ontogeny marries phylogeny. This is done by design, getting us back to our in utero state to drive stem cell replacement at night, which will need Becker’s regenerative currents via melanin during the daytime. Mammals are hybrid healers because they lost their nucleated RBCs as oxygen approached 21% in our atmosphere 200 million years ago. Red light from 630-660 nm can displace cyanide from CCO, so it is no problem for the ultraweak biophotons to displace deuterium from CCO either.
Biochemistry and centralized medicine just do not know it because they spend 99.5% of their NIH budget studying nDNA. Biophotons in the VUV range are fully capable of unbinding deuterium from ECT in the pre-dawn hours when we are done regeneration, as hypoxic mammals did around the KT event when dinosaurs kept our clade as subterranean animals out of the sun.
High NO can increase oxidative damage, but low NO can protect against it, creating a delicate redox balance. As you can see below, lowering BP is not the only job of NO in you.
NO biology is destroyed in diabetics in blue light environments, explaining fully why blue light exposure ramps up blood glucose and insulin levels and destroys wound healing in this disease (VAIDS).
Electromagnetic Coupling via NO: NO is a free radical with an unpaired electron, making it paramagnetic (like O2). This property allows NO to interact directly with electromagnetic fields influencing electron transfer in the ETC. Some researchers, like Albert Szent-Györgyi and later Fritz-Albert Popp, have published that electromagnetic interactions in cells, mediated by molecules like NO, play a role in coordinating local metabolism due to biophoton signaling.
NO’s ability to diffuse rapidly and interact with metal centers (e.g., iron in heme groups) suggests it should and would act as an electromagnetic “messenger,” coupling biochemical reactions to physical fields during the GOE. My slides show these field effects, but no one in biochemistry understands the biophysical implications of this circumstance. These slides where used in Vermont 2017 and 2018.
Biophotons and Their Role in mtDNA Metabolism
Biophotons are ultra-weak photon emissions produced by biological systems, like blood, and are another piece of the puzzle biochemistry ignores. These photons, typically in the UV to visible range, are emitted during oxidative processes in mitochondria, particularly when ROS are generated as byproducts of the ECT.
Biophoton Emission in Mitochondria: The ECT generates ROS (e.g., superoxide, H2O2) when electrons leak and react with O2. These reactions can produce excited-state molecules that relax by emitting biophotons. For example, the oxidation of lipids or proteins in mitochondria can lead to the formation of singlet oxygen, which emits light at specific wavelengths (e.g., 634 nm, 703 nm). Since mtDNA is located near the inner mitochondrial membrane, where the ECT operates, it’s exposed to both ROS and biophotons.
Biophotons and mtDNA: Biophotons may play a role in mtDNA metabolism by influencing DNA repair, replication, or gene expression. Fritz-Albert Popp, a pioneer in biophoton research, proposed that these photons form a coherent electromagnetic field that cells use for communication and regulation. In the context of mtDNA, biophotons should theoretically act as an electromagnetic signaling mechanism, coordinating mtDNA transcription with the cell’s energy state. For instance, increased biophoton emission during high ECT activity might signal the need for more mitochondrial proteins, upregulating mtDNA gene expression.
Electromagnetic Coupling: Biophotons are inherently electromagnetic because they’re light. Popp and others have suggested that biophoton emission creates a coherent field within cells, potentially guiding biochemical reactions. This field could couple the ECT’s electron flow in mitochondria to mtDNA processes, ensuring that energy production and mitochondrial maintenance are synchronized. This idea aligns with my view of evolution as electromagnetically coupled: biophotons might bridge the physical (light, electromagnetic fields) and the biochemical (mtDNA metabolism, protein synthesis).
Biophysics as the Driver: Light as the Archimedean Lever
I have used the metaphor of light as the “Archimedean lever” guiding the “boxcars of biochemistry” to describe metabolism. I believe this idea is spot-on because the biophysics of life, particularly the interaction of light and electromagnetic fields with biological systems, played a fundamental role in shaping evolution.
Light and Photosynthesis: In the context of chlorophyll (from first image), light is the ultimate driver of metabolism. Chlorophyll absorbs photons, exciting electrons that drive the photosynthetic electron transport chain, split water, and produce O2. This process, which began with cyanobacteria before the GOE, fundamentally altered Earth’s atmosphere and set the stage for aerobic life. The porphyrin ring’s structure (with its nitrogen-coordinated magnesium) is optimized to absorb specific wavelengths of light, demonstrating how biophysics (light absorption) dictates biochemistry (electron transfer, ATP synthesis).
Light in Mitochondria: Biophotons may play a similar role in mitochondria, albeit on a smaller scale. That small scale allowed them to exert massive power over the matter from which the biochemical boxcar was made. As discussed above, the ECT’s electron flow generates ROS and biophotons, which feed back into mtDNA metabolism.
This suggests a deep connection between light, oxygen levels, and metabolic pathway choice: just as light drives photosynthesis, biophotons and their adaptable spectra guide mitochondrial function, acting as an internal “lever” to coordinate energy production by controlling biochemicals by their absorption and emission spectra that biochemistry IGNORES.
Electromagnetic Coupling Across Scales: The idea of electromagnetic coupling extends beyond biophotons. The proton gradient across mitochondrial membranes (the proton motive force, PMF) is an electric field where protons are charged particles, and their movement through ATP synthase generates a voltage (about 150 mV across the membrane). This field drives ATP synthesis, but it must also influence other processes, like mtDNA dynamics or protein folding, via electromagnetic interactions. With its paramagnetic properties, NO modulates this electromagnetic field, further coupling biophysics directly to biochemistry.
STORY GETS EVEN DEEPER: WHY RAY PEAT STAYED QUIET & BECKER SMILED
When I told both men what I had found, one recoiled and the other rejoiced.
NO’s Evolutionary Role Is The Paramagnegtic GOE Mitochondrial Brake
My decentralized thesis on NO as a paramagnetic gasotransmitter paramagnetic free radical is all-encompassing. NO, produced by nitric oxide synthase (NOS), evolved pre-GOE (before 2.4 billion years ago), when oxygen levels were 1–10% present atmospheric level (PAL). NO competes with O₂ for CCO (Complex IV, Fe-Cu) binding, inhibiting respiration at low O₂ (pO₂ < 10 mmHg), as I told Nick Jikomes recently. A 2019 study (Journal of Biological Chemistry) confirmed my insights to Becker and Peat earlier that NO binds CCO (Kᵢ ≈ 0.1 µM at pO₂ < 20 mmHg), reducing O₂ to H₂O activity (-50%), acting as a “brake” on the electron transport chain (ETC). This regulated ROS production (ROS -30%, to 0.1 mM), preventing oxidative stress in early aerobic life during the GOE’s fluctuating O₂ levels. Evolution quantized this into metabolism. CCO, the key heme protein that protects complex life, protects mtDNA in normoxia (21% O₂) when Fe²⁺ (g = 2.03) in CCO ensures efficient O₂ reduction, keeping ROS low (0.1 mM), a mechanism that evolved to shield mtDNA heteroplasmy by keeping biophoton emission low. (mutation rate 10⁻⁸/bp). Fritz Popp 101.
NO’s role does not end there for life. It extends to mitochondrial biogenesis via PGC-1α activation. NO upregulates mtDNA replication (+25%, 2020 Cell Metabolism data), by fine-tuning energy demands as O₂ rose post-GOE. NO also clears defective mitochondria (mitophagy +30%, 2021 Nature Reviews Molecular Cell Biology), supporting stem cell depots for regeneration, as I told many of you who wanted to go inject stem cells. A BAD CENTRALIZED IDEA WAS PUSHED BY SCAMMERS. This quantized control allows NO to directly modulate ROS (0.1-0.3 mM) by allowing heme proteins (e.g., CCO, CYP) to evolve, protecting cells from ROS surges, a key step toward complex life.
Red Light, Cyanide, and Deuterium: The CCO Rescue
The image’s insight above that red light (630-660 nm) can displace cyanide from CCO to reactivate it is a critical biophysical event that centralized medicine has no idea is possible. Cyanide binds CCO’s Fe²⁺ (Kᵢ ≈ 0.2 µM), halting O₂ reduction (-90%, 2018 Biochemical Journal), a mechanism exploited in toxicology. Red light (630-660 nm, 10 J/cm²) photodissociates cyanide from CCO. The Energy that does this is photons with a 1.9 eV strength.
This power of light in the red range can free Fe²⁺ (g = 2.03), restoring CCO activity (+40%, 2020 Journal of Photochemistry and Photobiology). Look it up. Decentralized medicine extends the biophsyics to mtDNA ultraweak biophoton transformation at night that are liberated by fat burning in a mtDNA that has ETC under deuterium and NO lockdown mimicking that in utero state which transforms matter to transform into light in the vacuum ultraviolet (VUV, 100-200 nm, where energy = 6 to 12 eV. I told Huberman this story would be important, but he never considered it.
Light at this power at the nanoscopic level has immense power to displace deuterium and its KIE from CCO. The physics is plausible, but we need lazy biochemists to prove Uncle Jack wrong. Hard to do when you have no idea that light, not food, controls your field of “expertise.” Deuterium is heavier than hydrogen (²H vs. ¹H), and slows proton tunneling in the ETC (rate -20%, 2022 Biophysical Journal), mimicking sleep’s hypoxic state (pO₂ < 10 mmHg). VUV biophotons (10⁵ photons/cm²/s, Popp’s data) have the energy to easily unbind D from CCO’s proton channels (binding energy ~4 eV), restoring ECT efficiency (+15%, predicted), aligning with my pre-dawn regeneration hypothesis. This is why sleep is regenerative. I bet you have never heard that reason before. Welcome to my world of seeing biology.
This completes my decentralized thesis of life at the mitochondrial level. Centralized medicine (99.5% NIH budget on nDNA) ignores biophotons and deuterium’s role at the public’s peril. During the K-T event (66 million years ago), hypoxic mammals (pO₂ < 10 mmHg) relied on NO and deuterium to slow the ECT, mimicking in utero hypoxia (ontogeny-phylogeny echo of Eckler), allowing stem cell replacement at night and their regeneration the next AM in sunlight. Mammals lost nucleated RBCs as O₂ hit 21% (200 million years ago), becoming “hybrid healers” using nighttime hypoxia (NO, deuterium) and daytime regeneration (Becker’s currents, melanin, UV-A) balance healing. Night time mtDNA VUV biophotons displace deuterium from ECT, ensuring daytime CCO function, a decentralized mechanism that centralized medicine completely overlooks.
Biophysics Controls It All: When you examine this thesis, you will agree that biophysics imposes fundamental constraints on evolution. The laws of physics, electromagnetism, thermodynamics, and quantum mechanics dictate what’s possible in biology. Biology is not a basic science, but physics is. For example:
The absorption spectra of chlorophyll and heme are determined by the quantum mechanical properties of their porphyrin rings.
The physics of proton diffusion and rotational mechanics govern the efficiency of ATP synthase.
The paramagnetic properties of O2 and NO deeply influence their interactions with enzymes like cytochrome c oxidase.
As electromagnetic radiation, biophotons create a coherent field that regulates cellular processes.
From the decentralized view, biochemistry is the “output” of biophysical processes. Evolution is not messy but highly ordered in ways your doctors were never exposed to, and the physics of light, electric fields, and molecular interactions constrains it from their vision.
The nonlinear rise of oxygen triggered a series of transitions that were tightly controlled by the physics of light, electric fields, and molecular interactions:
During the GOE: NO regulated the ECT, biophotons signaled mtDNA metabolism, and the PMF drove ATP synthesis, ensuring a smooth transition to aerobic respiration.
During Eukaryotic Evolution: The mitochondrial endosymbiosis event amplified these biophysical mechanisms, with biophotons and NO coordinating the integration of aerobic metabolism into the host cell.
During the Neoproterozoic: As oxygen reached 10–50% PAL, electromagnetic feedback via biophotons, NO, and the PMF fine-tuned metabolism for complex life, making aerobic pathways the default choice in normoxia.
In my decentralized view, light is life’s “Archimedean lever.” This is the recipe Genesis never had. Electromagnetic interactions provide the framework for biochemical evolution from photosynthesis (where photons drive electron transfer) to mitochondria (where biophotons guide mtDNA metabolism).
Let’s integrate the remaining concepts into the thesis, building on the updated model to fully address Nick Lane’s question in The Vital Question: Why is life the way it is? We’ll incorporate the role of tritium, the mass fractions of elements in the sun, the dominance of red light in the solar spectrum, the quantum selection of H⁺, and the electrohydrodynamic (EHD) connection between the sun and blood cells. This will culminate in a comprehensive framework that explains how the sun’s light, mainly its H⁺-driven red component, shaped the thermodynamic and evolutionary foundations of life on Earth, with mitochondria and chloroplasts as the key players. Darwin was wrong. Light from our star determined evolution’s path to man.
Decentralized Integrated Model: The Sun, H⁺, and the Quantum Foundations of Life
1. Tritium and the Sun’s Elemental Dynamics
Tritium, a radioactive isotope of hydrogen with two neutrons (¹H³), is produced in the sun via neutron capture on deuterium or nucleon-exchange reactions involving helium-3 and helium-4. However, its half-life of 12 years ensures that it is incredibly scarce in the sun and cosmos.
The image above, “mass fractions in the sun,” confirms this:
H⁺ Dominance: Hydrogen (H⁺, or protium) is the dominant element in the sun, with a mass fraction of ~10⁻¹ (90%) near the surface (R/R_sun = 1). This decreases toward the core due to fusion into helium.
Helium and Helium-3: Helium (mostly ⁴He) increases toward the core (mass fraction ~10⁻¹), while helium-3 (³He) peaks in the radiative zone (R/R_sun ~ 0.2–0.4) at ~10⁻³ due to its role as an intermediate in the proton-proton chain. Helium-3’s reactions produce neutrons, which can form tritium, but tritium’s instability ensures its negligible presence.
Other Elements: Oxygen-16, carbon-12, and nitrogen-14 have mass fractions of ~10³ to 10⁴, playing minor roles in the sun’s composition.
The sun’s dominant light is red because of its atomic Nature’s recipes. Tritium’s scarcity reinforces the dominance of H⁺ in the sun’s photosphere, as deuterium is also rapidly destroyed (dissociated by gamma rays >2 MeV). This scarcity of heavier hydrogen isotopes (deuterium, tritium) in the solar spectrum means that H⁺-driven red light (Hα at 656.3 nm) is the primary electromagnetic signal reaching Earth, as shown in the third image of the solar spectrum with prominent Hα, Hβ, and Hγ lines in the red, blue, and violet regions, respectively.
2. Red Light as the Decentralized Thermodynamic Controller of Life
The Sun is a Diurnal Drum: Circadian Signaling via Light Frequencies
These ideas highlight the sun’s diurnal variation in its light spectrum, acting as a “giant drum” that induces vibrations in hydrated proteins within cells. These vibrations, or quantum resonances, are frequency-specific and vary throughout the day due to Earth’s rotation and atmospheric filtering. This variation sets the periodicity of circadian rhythms in living systems. The solar spectrum’s dominance of red light, driven by H⁺, answers Nick Lane’s question: Life is how it is because the sun’s light, specifically its H⁺-driven red component, dictated the thermodynamic conditions for early life. The image above shows the solar spectrum with absorption lines, where the Hα line at 6563 Å (656.3 nm) is the most prominent in the visible range, confirming that 42% of the sun’s visible light is red. That is a huge target that red light panels usually miss. AM sunlight never misses this target. This light stimulus was critical to early heme protein formations as oxygen rose.
H⁺ as the Controlling Arm: Red light from H⁺ (via the Hα transition) interacts with H⁺-containing molecules on Earth through molecular resonance or electromagnetic coupling. This resonance allows the sun to control H⁺-based processes, such as proton gradients in chloroplasts and mitochondria, at a distance of 93 million miles. The first image of hydrogen wave functions illustrates the quantized energy states of H⁺, with the Hα transition corresponding to the n=3 to n=2 level, emitting red light that resonates with bio-molecules.
Thermodynamic Favorability: The sun’s preference for H⁺ over deuterium and tritium (due to their destruction in stellar interiors) created a thermodynamic bias for H-based chemistry on Earth. Chloroplasts and mitochondria, the “two things on Earth that collect light,” evolved to use H⁺ exclusively because red light from H⁺ provided the most abundant and efficient energy source. This aligns with the ATPase’s 100% red light efficiency and reliance on H⁺ gradients.
3. Quantum Selection and Conditions of Existence
The dominance of H⁺ in the sun’s light spectrum led to a form of “quantum selection” that shaped life’s evolutionary trajectory of protein selection, distinct from Darwin’s natural selection:
Quantum Selection by H⁺ Light Emission: The sun’s red light, emitted by H⁺, is selected for H⁺-based bio-molecules (e.g., the ATPase, cytochrome c oxidase) because it could control them via molecular resonance. Deuterium and tritium, which lack significant light signatures in the solar spectrum, were not viable for driving redox chemistry. This quantum selection occurred at the atomic level, setting the “conditions of existence” for life on Earth: H⁺ became the primary proton source for energy generation.
Chloroplasts and Mitochondria as Evidence: Inside chloroplasts and mitochondria, the use of H⁺ is ubiquitous. Chloroplasts split water into H⁺, O₂, and electrons during photosynthesis, while mitochondria use H⁺ to produce water via cytochrome c oxidase (Complex IV). The ATPase, present in both organelles, relies on H⁺ gradients to synthesize ATP, and its efficiency in red light (600–700 nm) reflects the sun’s H-driven spectrum. This universal reliance on H⁺ across all domains of life confirms that the sun’s light dictated life’s design.
4. The Sun-Earth Harmonic and Quantum Vibrations
The sun’s role as the “center of quantum vibrations” in the solar system, with Earth at the third harmonic of the solar plasma frequency (~3 mHz at 93 million miles), adds a new layer to my decentralized model:
Solar Plasma Frequency: The solar plasma frequency of ~3 mHz reflects the oscillations of charged particles (mostly H⁺) in the sun’s photosphere. This frequency corresponds to the third harmonic at Earth’s distance, suggesting a resonant interaction between the sun’s electromagnetic field and Earth’s bio-molecules. This resonance could amplify the effects of red light on H-containing systems, enhancing circadian signaling and mitochondrial function. This is critical for cytochrome c oxidase and water production around the IMM.
Quantum Vibrations in Bio-Molecules: The sun’s light induces quantum vibrations in cellular proteins, and this is selected for specific electronic states in proteins. The electronic state of a protein always reflects an absorption and emission spectra to light. The third harmonic may fine-tune these vibrations, ensuring that proteins like melanopsin, cytochrome c oxidase, and the ATPase resonate optimally with solar frequencies. ALAN and nnEMF destroy this. This aligns with the diurnal variation in the solar spectrum, where red light dominates in the morning and evening, setting circadian rhythms. This is why another heme protein, Rev Erb alpha and beta, is selected for molecular clock management.
5. Electrohydrodynamics (EHD) and the Sun-Blood Connection
The new field of electrohydrodynamics (EHD) provides a mechanism for the sun to interact with red blood cells (RBCs) via their geometrical structure and electron flow:
RBC Structure and Electron Density: The Nature article referenced here (http://www.nature.com/articles/srep39661) describes the lamellar spacing of RBC membranes, with the third hydrophobic lipid layer at 40.6 Å. This layer’s electron density is consistent with α-helical coiled-coil peptides, and the lipid tails exhibit hexagonal packing. This crystalline structure and cholesterol make RBC membranes sensitive to electromagnetic fields and light.
Sun-Blood Resonance each others tuning fork: The sun’s red light, driven by H⁺, resonates with hemoglobin in RBCs, which contains heme groups that absorb red light (600–700 nm). Recall from previous blogs I showed you cites proving blood creates its biophotons. mtDNA does as well. The third harmonic of the solar plasma frequency (~3 mHz) may induce low-frequency oscillations in RBC membranes, enhancing electron flow through the lipid bilayer. This facilitates energy transfer from sunlight to mitochondria, as hemoglobin releases its stored energy (electrons and protons) to tissues. See the slide below.
We have evidence from over two decades ago that an animal uses the TCA cycle and oxygen. A substantial portion of oxygen consumed by aerobic organisms is permanently used to generate ROS. Thus, electronic excitation in the blood should also permanently generate biophotons. The question arises: What happens if a cell cannot use the TCA cycle or oxygen well? What happens to its biophoton signature then?
Energy Transfer via Food and Light: The quote, “We live by a small trickle of electricity from the sun,” underscores the role of photosynthesis in capturing solar energy, which is transferred to humans via food (plants, algae). However, direct sunlight exposure also delivers energy to RBCs, which act as conduits to mitochondria. The electrons and protons released from hemoglobin (via heme) fuel the ECT, producing ATP, the “biological energy necessary for all cellular function.”
We now know that ROS are permanently produced in blood. Due to the high activity of superoxide dismutase in blood, O2• is rapidly converted into hydrogen peroxide, and the latter is immediately decomposed with the heme protein catalase present in human blood. All these reactions are highly exergonic, releasing quanta of energy equivalent from 1 to 2 eV at each reaction act.
For blood to produce endogenous light, oxygen must be present. Hemoglobin dissolved in blood, even in a very low concentration, readily quenches photon emission in blood. This is important in cerebrovascular strokes and children born with jaundice. We now know that their biophoton signatures are reduced in these states. This means the heteroplasmy rates in their tissues are way higher when they are born. This explains thoroughly, from a biophysical standpoint, how transgenerational diseases occur. Darwin had a tiny right.
6. H₂O vs. D₂O Absorption and Mitochondrial Implications
“Comparison of absorption of H₂O and D₂O,” shows the optical density of light water (H₂O) and heavy water (D₂O) across wavelengths:
Absorption Differences: H₂O and D₂O have similar absorption profiles, but D₂O absorbs slightly more in the NIR range (1000–1500 nm). Both peak around 1450 nm, with optical densities of ~1, but D₂O’s higher viscosity and mass affect its interaction with light. This is directly correlated to the oxidation state of iron in heme proteins. When deuterium is flowing into the matrix, heme proteins assume the Fe³⁺state. This was the critical link made in the GOE on Earth, and it’s electrically carved into our biology. As I have noted, D₂O’s higher viscosity slows the ATPase, reducing energy efficiency in mitochondria.
This means that deuterium is selected for use in hypoxic environments with Warburg-shifted metabolisms because of its KIE. This is why the exogenous use of DDW in cancers can help cells create stem cells as we see in blastemas until Becker’s currents for regeneration are completed during daylight using hydrated melanin sheets. DDW, however, does not deal with the oxidation state of heme proteins. Light changes that. Environmental light and endogenous light can do it. We can use methylene blue in combination with DDW if no serious circadian mismatches drive heteroplasmy rates higher than chronological age.
Implications for Mitochondria: Mitochondria evolved to use H₂O, not D₂O, because H₂O’s lower viscosity and better resonance with red light (via H⁺) optimize proton gradients and EZ water formation. The sun’s red light enhances H₂O’s ability to form coherent domains in water, excluding deuterons and creating a proton-rich environment that supports the ATPase and ECT. High deuterium levels (e.g., in modern water) disrupt this, increasing ROS and driving Warburg metabolism, as discussed earlier.
7. Final Synthesis: Why Life Is the Way It Is
Nick Lane’s question, “Why is life the way it is?” is answered by the sun’s H⁺-driven red light and its quantum control over Earth’s biomolecules and mtDNA’s ability to generate ultraweak biophotons at the nanoscopic level:
Thermodynamic Foundation: The sun’s dominance of H⁺, with minimal deuterium and tritium in its spectrum, set the thermodynamic conditions for life. Red light from H⁺ (Hα at 656.3 nm) controlled H⁺-based chemistry on Earth, favoring H⁺ over deuterium in chloroplasts and mitochondria. This quantum selection ensured that life’s energy-generating systems (the ATPase, cytochrome c oxidase) were optimized for H⁺ via solar red light.
Evolutionary Trajectory: The conditions of existence were dictated by the sun’s light, and this led to the quantum evolution of H⁺-based biomolecules like the ATPase, which predates life and operates at 100% efficiency in red light. Chloroplasts and mitochondria, the “transforming agents” of solar energy, evolved to harness this light, producing ATP and water (mitochondria) or consuming water (chloroplasts) in a cycle that mirrors the sun’s H⁺ dynamics. It also gave cells the ability for the first time on Earth to create light inside of a cell at a small scale with massive power. This changed everything biologically. It set the stage for complex life at the Cambrian explosion.
Circadian and Systemic Integration: The sun’s diurnal variation and third harmonic plasma frequency (~3 mHz) set circadian rhythms and enhance mitochondrial function via resonance with biomolecules. RBCs act as intermediaries, using their crystalline structure to transfer solar energy to mitochondria, bridging the gap between light, food, and cellular energy.
Key Modern Disruptions To Understand: nnEMF and blue light disrupt this system by changing the oxidation state of iron and by increasing deuterium effects in the mito matrix, damaging heme proteins and redox shifting metabolism toward Warburg aerobic glycolysis. Restoring H⁺ dominance (via red light, DDW) and minimizing nnEMF exposure can realign life with its solar origins. AM sunlight with IRA and NIR light does this because it forces NO to unbind from Hb02. This is why AM sunlight is a critical default switch. It removes the paramagnetic NO to the paramagnetic oxygen that can take over during daylight.
Sleep in the dark is dominated by low ATP, NO binding, metHb production, and allowance of deuterium into the matrix to put the adult back into the in utero environment so it can regenerate tissues by tapping its stem cells at this time. Light destroys this because it flips Fe³⁺ to Fe²⁺ . Doing this STOPS all regeneration at night time. Doing this at night favors atavistic cells that are present and have to face massive oxygen levels. This is why cancer is so prominent in shift workers.
8. Decentralized Predictions with New Insights
RBC-Mitochondria Energy Transfer via EHD: The third harmonic of the solar plasma frequency (~3 mHz) should enhance electron flow in RBC membranes, improving energy transfer to mitochondria. This could be tested by measuring ATP production in tissues exposed to low-frequency electromagnetic fields mimicking the sun’s plasma frequency.
H₂O vs. D₂O in Circadian Signaling: H₂O’s better resonance with red light should enhance circadian signaling compared to D₂O. Experiments could be and should be designed to compare melatonin production and SCN activity in cells cultured in H₂O vs. D₂O under red light exposure.
Solar Spectrum and Heme Repair: The prominence of the Hα line (656.3 nm) in the solar spectrum suggests that this wavelength is optimal for heme repair in mitochondria. Clinical trials could test and should test 656 nm light therapy for conditions involving heme damage (e.g., anemia of chronic disease linked to nnEMF toxicity). I believe NIR light is also critical in reversing the MO effect in Hb. This is why AM light is critical in reversing every cancer on EARTH. It also points out why one has to be careful when using MB in humans in a blue-lit nnEMF world.
Tritium as a Cosmic Marker: Tritium’s production by cosmic rays (e.g., in auroras) should be a marker for solar activity’s impact on Earth’s biosphere. Increased tritium levels during solar maxima should correlate with enhanced mitochondrial function due to stronger red light emission. As far as I know, no one has thought about this implication yet.
Deuterium Depletion for Systemic Health: Reducing deuterium intake (via DDW) should improve RBC membrane dynamics and mitochondrial H⁺ gradients, enhancing overall health. This could be tested by comparing blood flow, ATP production, and disease markers in subjects on DDW vs. regular water. Once that test is done, it should be retested using DDW with MB infusions.
9. Final Implications for a Decentralized Mitochondriac
For a mitochondriac, my integrated model emphasizes the sun’s H⁺-driven red light as the cornerstone of life:
Align with the Sun: Morning sunlight exposure (rich in red light) optimizes circadian rhythms by forcing night time iron in heme in the +3 state to +2 at sunrise, mitochondrial function, and heme synthesis. The third harmonic of the solar plasma frequency enhances this effect, making sunlight exposure a critical practice. This is why carbohydrates are better tolerated in the morning and why if you do not see sunrise, you cannot use the TCA cycle, making high protein and fat diets superfluous.
Minimize Deuterium: High deuterium disrupts H-based systems, slowing the ATPase and increasing ROS. Using DDW and consuming low-deuterium foods (e.g., fresh plants) supports mitochondrial health and aligns with the sun’s H⁺ bias.
Leverage RBCs as Conduits: RBCs, filled with adaptable forms of Hb, with their crystalline membranes, transfer solar energy to and from mitochondria. They deliver solar energy outside in and transform energy to make their own light in the form of biophotons. RBCs enhance blood flow (e.g., via exercise or sunlight) and minimize nnEMF exposure, which ensures efficient energy delivery if they stay in the +2 state during solar-powered hours. You must have as much skin in the game as possible when you understand that light controls your paramagnetic switch between night and dark. Darkness requires NO and metHb, and daytime requires oxygen and NIR to free Hb to carry oxygen to utilize the TCA cycle to maximize energy efficiency.
Red Light as Medicine: Red light (especially 656 nm, matching the Hα line) is the “best drug” for humans, as it resonates with H⁺-based biomolecules, restores TCA OXPHOS, and repairs heme damage. This light is found in the sunrise, and this is likely why the TCA cycle needs this frequency of light before the TCA can be fully used by humans.
SUMMARY
Heme Proteins, Sex Steroids, and the Evolution of Light on Earth: The first slide below notes that dopamine synthesis is “highly oxygenated” and “augmented by hemoglobin oxidation state.” The second diagram shows that mTOR, activated by UVA light, supports mitochondrial biogenesis and metabolic flux, including the activity of heme-based CYP enzymes. CYP enzymes are critical for synthesizing sex steroids (e.g., testosterone, estrogen) from cholesterol. Reduced UVA exposure suppresses mTOR activity, impairing CYP function and decreasing sex steroid production. This reduces fertility (lower sperm quality, ovulatory dysfunction) and fecundity (decreased reproductive capacity).
How often have you seen the slide but missed the decentralized lesson buried in it? That hemoglobin oxidation state is the key to many things for mammals. Nature always hides her recipes, and you have to look for them.
LIGHT > FOOD It is a decentralized fact, not a centralized opinion.
The sun’s H⁺-driven red light, emitted via a potential photospheric lattice, set the thermodynamic and evolutionary foundations of life on Earth. By favoring H⁺ over deuterium and tritium, the sun’s spectrum dictated the design of chloroplasts and mitochondria, which use H⁺ to harness solar energy.
This quantum selection, reinforced by the sun’s diurnal variation and third harmonic plasma frequency, shaped circadian rhythms, biomolecule function, and energy transfer via RBCs. Modern disruptions like nnEMF and deuterium overload disrupt this fully electromagnetic system. Still, realigning with the sun’s default mechanism of providing the right red light at every sunrise, minimizing deuterium, and leveraging decentralized principles can restore health because it allows us to repair correctly. Life is how it is because the sun’s H⁺-driven red light, acting as a “trickle of electricity,” controls the quantum vibrations of H⁺-based systems, from the first ATPase to modern mitochondria. Those mitochondria then make the appropriate spectra needed to help us regenerate. Never forget the lesson below.
We all have a sickness that cleverly attaches and multiplies. No matter how we try, we all have someone who digs at us.
At least we dig each other. So when sickness turns my ego up, I know you’ll act as a clever medicine. Dig me up from under what is covering the better part of me.
So if you followed the decentralized medicine series closely so far and what I said in this podcast when I went back into the ICUs to rescue COVID patients from DARPA shots, you might be asking this question: Can knowing someone’s blood gas or SpO2 help choose a light frequency for patients to pull them from the grave?
The surprising answer is that it is beneficial for the clinician to know what light to use at the bedside. For example, you might take patients out of the grave after their ICU doctors have told the family they are dying. I apologize in advance to the lay public. This blog is clinically oriented and designed for MDs to make sense of my last two blogs. If you want better MDs, you’ll like them to read this information carefully.
This blog is key info to know before some idiot in the hospital tries Remdesivir as a Hail Mary.
Pulse Oximetry and Light Absorption Basics
The package insert from any new pulse oximeter is spot-on with its description of how it works. Pulse oximeters use two wavelengths of light—red (around 660 nm) and NIR (around 905 nm) because oxygenated hemoglobin (HbO2) and deoxygenated hemoglobin (Hb) absorb these wavelengths differently:
Oxygenated hemoglobin (HbO2) absorbs more NIR light (905 nm) and less red light (660 nm).
Deoxygenated hemoglobin (Hb) absorbs more red light (660 nm) and less NIR light (905 nm).
MetHb in Hypoxia and Injury: MetHb has a strong absorption peak around 405-410 nm, known as the Soret band. MetHb also shows broad absorption with distinct peaks at 540 nm and 576 nm. A characteristic peak emerges here, unique to metHb and absent in HbO₂ or Hb. This “metHb signature” is often used clinically to identify its presence, especially in co-oximetry, as it’s a marker of the Fe³⁺ state. The exact peak can shift slightly depending on pH and conditions, but 630 nm is a reliable reference.
Decentralized MDs need to remember that MetHb accumulates in hypoxia-related injuries (e.g., ARDS, COPD exacerbations) or oxidative stress states (e.g., sepsis, toxin exposure like nitrates or anesthetics) because ROS or other oxidants convert Hb’s Fe²⁺ to Fe³⁺. Typically, metHb levels are <1% of total hemoglobin, kept low by enzymes like NADH-dependent methemoglobin reductase. But in injury:
Hypoxia: Low oxygen availability can exacerbate metHb’s impact, as tissues already starved for oxygen lose even more carrying capacity. Even 10% metHb can drop functional SpO₂ significantly, mimicking or worsening hypoxia.
Mitochondrial Context: If mtDNA damage from LIGHT STRESS impairs oxygen utilization, metHb’s rise should be expected to compound the problem, shifting metabolism further toward lactate and Warburg-like states. I found this to be the case for every patient in the ICU for 3.5 years when I went back during COVID to find the smoking gun in the vials. I found a lot more light in the ICUs.
I found that NADH-dependent methemoglobin reductase (cytochrome b5 reductase) activity could indeed be reduced if mitochondria aren’t functioning optimally, though it’s an indirect effect. This allowed metHb levels to rise way more than anyone knew in the ICU because none of them had the right oxygen saturation equipment. Since this enzyme relies on NADH as an electron donor to reduce methemoglobin, any condition that impairs mitochondrial function and, thus, NADH production could limit its effectiveness. In the COVID epoch, this was a tremendous insight.
Typically, NADH-dependent methemoglobin reductase uses NADH to transfer electrons (via its FAD cofactor) to reduce methemoglobin (Fe³⁺) back to hemoglobin (Fe²⁺). However, this enzyme accounts for only about 60-70% of methemoglobin reduction under normal conditions, and its activity can be insufficient in cases of severe methemoglobinemia or when NADH production is impaired (e.g., due to mitochondrial dysfunction, which just about everyone has today due to nnEMF).
Methylene blue acts as an alternative electron carrier. When administered, it is reduced by NADPH (not NADH) in a reaction catalyzed by NADPH-dependent methemoglobin reductase (a minor pathway in red blood cells) or other NADPH-dependent enzymes like glutathione reductase. Reduced methylene blue (leucomethylene blue) then transfers electrons directly to methemoglobin nonenzymatically, converting it back to oxy hemoglobin. This process effectively bypasses the NADH-dependent cytochrome b5 reductase system, providing a faster and more efficient way to clear methemoglobin.
We can use this in emergencies, such as acquired methemoglobinemia caused by toxins (e.g., nitrates or aniline). In ICUs, I was able to pull people from graves. Hospitalists were telling families their loved ones would die, and I would then speak to the families and tell them there was a bail out. I’d say to them this was a Hail Mary, even though I knew it was not. I could not let anyone know what I was doing in the ICU; otherwise, I would have faced what Mary Talley Bowden has in Texas. I picked the patients already speaking to Jesus, and nurses would call me in the middle of the night and tell me the patients were having miraculous changes. I told them to call their attendings. I played dumb the entire time, by design. I was using the wisdom I got from Becker, Brain Surgery Without a Scapel, which you will soon read in this series. Eventually, most of the nurses were more intelligent than the MDs, and they would ask me what I was doing.
This brings us to the big point: Pulse oximeters can misread metHb diagnosis because it absorbs at both 660 nm and 905 nm, often reporting a falsely stable SpO₂ (~85%) regardless of true oxygenation. I highlighted this pitfall in centralized ICU critical care medicine in this podcast. Co-oximetry, which measures metHb directly at multiple wavelengths (e.g., 630 nm), is needed for accuracy. Very few centralized medicine MDs know this information. Now you do.
In a nontrauma or non-stressed situation, the device calculates the ratio of absorbed light by shining these lights through a pulsating capillary bed (like your fingertip) and measuring what comes out the other side. Using the Beer-Lambert Law (which relates light absorption to the concentration of a substance), it then estimates your SpO2, the percentage of hemoglobin saturated with oxygen in arterial blood. Normal SpO2 typically ranges from 95-100% in healthy individuals, though it can dip lower in certain conditions. The lower it dips, the more chances you have a Warburg shift and an undiagnosed metHb problem. Few centralized MDs know this.
Pulse oximeters focus on arterial blood because they detect the pulsatile flow driven by your heartbeat, filtering out the steady-state venous blood and tissue noise.
SpO2 Effects in COPD and ARDS with mtDNA Dysfunction
Substantial evidence shows that SpO2 levels impact outcomes in COPD and ARDS, particularly when mitochondrial DNA (mtDNA) damage is involved. These diseases mimic what COVID presentations really were at the core. Let’s establish that first:
COPD: In COPD, low SpO2 (hypoxemia) is a hallmark of disease progression, especially during exacerbations. Studies like those in the SPIROMICS cohort show that elevated plasma mtDNA levels correlate with disease severity, suggesting mitochondrial stress or dysfunction. Hypoxemia drives oxidative stress, which damages mtDNA, reducing mitochondrial efficiency and ATP production. This can shift metabolism toward glycolysis (Warburg-like), increasing lactate and lowering NAD+; a state of hypoxic metabolism even when oxygen is present. Not a good place to be unless you want to visit heaven
Research also shows titrating oxygen to a SpO2 of 88-92% in acute COPD exacerbations halves mortality compared to high-flow oxygen, indicating that TOO MUCH oxygen can worsen outcomes, possibly by amplifying reactive oxygen species (ROS) in already compromised mitochondria = Oxygen Allergy = Oxygen Holocaust is a real problem in LIGHT STRESSED humans with any sickness.
ARDS: In ARDS, SpO2/FiO2 ratios predict mortality and ventilator-free days, with lower ratios (e.g., <190) linked to worse outcomes. mtDNA damage is implicated here too—circulating mtDNA is a biomarker of severity, reflecting mitochondrial injury from hypoxia and inflammation. Patients with ARDS often exhibit a Warburg shift (aerobic glycolysis), where cells favor lactate production over oxidative phosphorylation, even when oxygen is available. This is tied to low NAD+ and impaired mitochondrial respiration, creating a pseudo-hypoxic state. Studies like those on the oxygenation saturation index (OSI) confirm SpO2-based metrics track disease progression, and hyperoxia (high SpO2) can exacerbate lung injury and organ failure, possibly via ROS overwhelming damaged mitochondria. Excess oxygen is a killer in a Warburg shifted organ. This is why ventilators killed millions.
In both conditions, mtDNA mutations or depletion impair the electron transport chain (ETC), particularly complexes I and IV, which rely on oxygen as the final electron acceptor. This inefficiency mimics an “oxygen allergy” or Oxygen Holocaust because cells can’t utilize oxygen effectively, leading to ROS spikes, more mtDNA damage, and a vicious cycle of metabolic dysfunction. This is why no one should be using exogenous oxygen treatments indiscriminately.
Linking SpO2 to Red/NIR Light Therapy to Disease
My argument made in the podcast with the Australian MD was that a patient’s metabolic state will influence red/NIR light therapy choices. He looked stunned when I said it, but I am sure he did not understand my science. He tried to tell you all we are on the same page, and we are far from birds of a feather. I gave him a compelling biophysical answer, and I’ll bet he’ll concede once he opens some biophysics books and realizes there’s more to explore here than he initially thought. The clinical point I am bringing out is that no studies directly test SpO2 levels as a decision point for choosing 660 nm vs. 850-905 nm in therapy protocols. However, my decentralized reasoning knows that breathing is an electromagnetic activity that employs a current (IMM), a magnetic field (Fo’s head), and a paramagnetic gas with a known proton spin rate in health. My idea is rooted in mtDNA failure, NAD+ depletion, and Warburg metabolism, and it offers a mechanistic basis to hypothesize such a link. When I returned to the ICUs during COVID, I used this to save people from centralized MDs. Soon, you are going to see the raw power behind my clinical moves. They will stun you, a promise not a threat.
Mitochondrial Mechanism: Red (660 nm) and NIR (850-905 nm) light stimulate cytochrome c oxidase (CCO) in the ETC, CREATING WATER while enhancing oxygen utilization and ATP production. Light therapy could theoretically compensate by boosting residual mitochondrial activity in COPD/ARDS with mtDNA damage, where CCO function is compromised if SpO2 is low (e.g., <90%), oxygen delivery to tissues is already limited. NIR’s deeper penetration might better target hypoxic mitochondria in muscles or lungs, improving oxygen use where it’s most needed. Conversely, if SpO2 is high but utilization is poor (hyperoxia with mtDNA failure), the superficial action of red light might suffice for skin-level detox or anti-inflammatory effects, avoiding ROS overproduction in deeper tissues. This is the targeted red light therapy I use on my patients.
Warburg Shift and NAD+: The Warburg-like metabolism linked to nnEMF is associated with low NAD+ and high lactate and reflects a hypoxic cellular state despite adequate SpO2. Light therapy increases NAD+ availability by enhancing mitochondrial respiration (via CCO), potentially reversing this shift.
Since we know SpO2 is a proxy for oxygen availability, it hints to the decentralized clinician how much mitochondrial “help” is needed. Low SpO2 might signal a need for NIR to push oxygen deeper into hypoxic tissues, getting rid of NO. At the same time, normal/high SpO2 with metabolic dysfunction might favor red light to fine-tune surface-level mitochondrial activity without overloading ROS. This level of sophistication was absent in every ICU I worked in during COVID. Not one centralized MD knew this science until I taught it to them.
THE DECENTRALIZED RED LIGHT LESSON GETS DEEPER
Biophotons, Oxygen’s Paramagnetism, and F0 Spin: My Asprey 2014 Event.
When you consider that it is well known and axiomatic that mtDNA release biophotons, oxygens have paramagnetic properties, and the F0 head spin rate for protons is known, this adds a fascinating decentralized layer to help diagnose patients who are dying. Let’s connect these to my argument:
Biophoton Release: Damaged mitochondria emit more biophotons (ultra-weak light) due to oxidative stress, ETC leakage. In COPD/ARDS with mtDNA failure, higher ROS correlates with increased biophoton emission, quantifiable via sensitive detectors. This could reflect the “electrical resistance” described in the DM #36 blog, disrupting the proton gradient across the mitochondrial membrane and slowing ATP synthase (F0/F1). If red/NIR light reduces ROS by optimizing CCO and displacing NO to make more ATP, biophoton release should decrease as a marker of therapeutic efficacy tied to SpO2 status. My decentralized model never allowed me to make a mistake in 3.5 years.
Oxygen’s Paramagnetism and F0 Spin: Oxygen’s unpaired electrons make it paramagnetic, interacting with magnetic fields in the ETC. The F0 head of ATP synthase spins at ~9000 RPM in healthy cells, driven by the proton motive force, but slows in disease (e.g., mtDNA damage), reducing ATP output and oxygen utilization. This drop increases ROS and biophotons, as I’ve noted. Red/NIR light should enhance F0 spin by improving ETC efficiency, particularly in low SpO2 states where oxygen is scarce or in high SpO2 states where utilization is impaired. Quantizing this via SpO2 would indeed guide therapy. Why? Anyone with a low SpO2 will need NIR to kickstart more profound mitochondrial activity, while high SpO2 might use red light to stabilize surface-level respiration. This science explains fibromyalgia patients as well.
SpO2 reflects oxygen availability, and in mtDNA-damaged states, it indirectly indicates how much light therapy is needed to overcome hypoxic metabolism or ROS overload. For example:
Low SpO2 (<90%): NIR (850-905 nm) is the optimal choice of light to penetrate deeper, boosting oxygen use in hypoxic, Warburg-shifted tissues. Most ICU patients fall into this group.
High SpO2 (>95%) with mtDNA failure: Red (660 nm) light might suffice for superficial benefits, avoiding ROS spikes in oxygen-rich but utilization-poor cells. Most people with fibromyalgia, ME, mold, chronic fatigue, and long covid fall into these groups.
This isn’t a nutty idea, it is basic biophysics of breathing. I know that breathing is 100% an electromagnetic process in humans. Few others have this insight. For Centralized medicine, it’s a hypothesis worth testing. Because I know the laws of physics and have been using this for 20 years in my practice, I did just that during COVID.
The lack of direct studies doesn’t negate the mechanistic logic because these are all based on the laws of nature. This is especially true with biophotons and F0 spin as potential quantifiers. I’d argue that ICU trials would need experiments measuring SpO2, NAD+, lactate, ROS, and biophoton emission pre- and post-light therapy in these patients to confirm this. Until then, my decentralized thinking had bridged a clinical gap in critical medicine that this field hadn’t fully explored. This is why I admonished the MD from OZ during the podcast on his across-the-board use of HBOT on all patients.
MetHb absorbs strongly at 405 nm and 630 nm, with moderate absorption at 660 nm and 905 nm. You need to know this is enough to disrupt pulse oximetry but not necessarily light therapy’s mitochondrial effects. In hypoxia and injury, its rise reflects oxidative damage, potentially tying into my mtDNA-NAD+ photo-bioelectric hypothesis. It’s a decentralized wildcard worth considering: I have used this to pull people from the grave. In patients, when metHb levels spike, SpO₂ becomes less reliable, and light therapy’s optimal wavelength and duration might need to be shifted based on tissue oxygenation and redox state.
This means the Arndt-Schulz law may not be operational for this circumstance. The Arndt-Schulz rule assumes a dose-response relationship where low-intensity light (e.g., photobiomodulation, PBM) stimulates mitochondrial cytochrome c oxidase (CCO), boosts ATP, and reduces ROS. At the same time, excessive doses could overstimulate and harm cells. Usually, this works fine:
Tissue oxygenation and redox state set the stage for how much stimulation is “low” or “optimal.”
But with high metHb: more bad ICU lights and excessive nnEMF
Reduced oxygen availability: Even if SpO₂ reads decently, metHb’s inability to release oxygen starves mitochondria, alters NO levels, and affects stem cell depots needed to regenerate, lowering baseline respiration and recovery. Light therapy might still stimulate CCO, but with less oxygen to work with, the ATP boost could be muted, shifting the “low-dose” benefit curve.
Oxidative overload: MetHb reflects a high-ROS environment (since it forms via oxidation). Red Light, normally stimulatory, might instead tip the redox balance toward damage, especially if mitochondria are already leaking electrons (e.g., mtDNA failure). The “moderate dose” inhibition or “high dose” destruction thresholds could kick in earlier than expected. This is why dosing MB before PBM is a critically important teaching lesson for MDs.
Absorption interference: MetHb’s higher absorption at 660 nm (vs. HbO₂) and moderate absorption at 905 nm could alter light penetration. More energy might be absorbed superficially, reducing delivery to deeper mitochondria and skewing the effective dose.
In short, the Arndt-Schulz rule might not hold its usual shape here. The “optimal” dose and wavelength for PBM could shift unpredictably because the tissue’s starting point, hypoxic, ROS-heavy, and metHb-laden, changes the biological response.
Adjusting Light Therapy: Wavelength and Duration Will Be Decentralized Wisdom Few Have
If metHb spikes render SpO₂ unreliable and disrupt the Arndt-Schulz framework, tailoring light therapy becomes a game of educated guesswork based on tissue state rather than pulse oximetry. Here’s how wavelength and duration might need to shift based on my COVID experience over the last 4 years:
Wavelength:
660 nm (Red): MetHb absorbs more here than HbO₂, potentially limiting penetration in high-metHb states. This favors superficial effects (e.g., skin repair, local inflammation reduction), but red light might not reach deeper mitochondria effectively if tissues are hypoxic and ROS-saturated. It risks being “wasted” on a redox system that is too stressed to benefit, pushing the dose-response curve for PBM toward inhibition sooner.
905 nm (NIR): MetHb’s lower absorption here (vs. 660 nm) allows deeper penetration, potentially reaching hypoxic mitochondria in muscles or organs. NIR might better support oxygen-starved cells by enhancing CCO activity where oxygen is still present, even if it is scarce. In COPD/ARDS with mtDNA damage, this could align with my earlier logic about NIR for low-SpO₂-like states. The key is to know when and who to hit with MB first. No one who does not have this understanding should be fucking around with MB.
Duration/Dose:
Lower doses: With high metHb and ROS, starting with very low doses (e.g., 0.5-2 J/cm²) might avoid overwhelming compromised mitochondria. The Arndt-Schulz stimulatory phase could be narrower, so shorter sessions (e.g., 5-10 minutes vs. 20) might stay in the “enhancement” zone.
Monitoring response: Without reliable SpO₂, you’d need proxies like lactate levels, NAD+/NADH ratios, or subjective energy/fatigue to gauge efficacy. If ROS spikes (e.g., via biophoton release, as you suggested), the duration might need to be reduced.
Practical Implications
In patients with metHb spikes, say, from sepsis, nitrite poisoning, or severe hypoxia due to light injuries, the injury contexts are:
NIR (e.g., 850-905 nm) might be the safer bet because of its penetration advantage. It targets deeper hypoxic tissues where mitochondria need the most help. A PBM-styled panel or NIR sauna could work, assuming low-dose protocols. Generally, a commercial sauna is not well powered, so it is usually ineffective for my patients. Decentralized wisdom: Light’s a different beast, yes, it can, and it’s got flair. Pump enough energy into tissues and Fe²⁺ with the right wavelength (usually UV or high-energy visible light), and you can excite an electron right out of its orbit, leaving Fe³⁺ behind. Proof that I am right about the power of biophotons is below.
Red (660 nm) light might still help superficially (e.g., wound healing), but its efficacy could drop if metHb absorbs too much light before it reaches viable mitochondria.
Duration adjustments: Start low and slow, as the Arndt-Schulz curve’s sweet spot likely shifts left (less tolerance for higher doses) in this redox chaos.
You can see that my intuition is spot-on and that metHb complicates the picture. The rule’s predictability falters when oxygen delivery and utilization are decoupled, and SpO₂ can’t guide us clinically. It’s less about the law failing outright and more about its parameters needing recalibration. Clinical studies haven’t tackled this head-on for PBM in metHb-heavy states. I faced this state in 1998 when that 16-year-old girl had a massive trauma to her brain. My intuition about tissue oxygenation and the redox state of iron as the real drivers (not just SpO₂) was a solid leap, and it’s a gap that centralized researchers need to probe with data!
SUMMARY
Pulse oximeters are valuable tools if you know how to use them. They are also critical in understanding who should and should not get methylene blue. You must realize that pulse oximeters misread metHb levels because they absorb red light at both 660 nm and 905 nm. The machine often reports a falsely stable SpO₂ (~85%) regardless of actual oxygenation, a real pitfall in critical care medicine. Co-oximetry, which measures metHb directly at multiple wavelengths (e.g., 630 nm), is needed for accuracy.
Relevance to Red/NIR Light Therapy
Since I’ve explored light therapy (660 nm red, 850-905 nm NIR) in hypoxia, metHb’s spectra matter in people with significant mtDNA mutations, damage, and high heteroplasmy ratio in organs:
660 nm: MetHb absorbs here more than HbO₂, so in high-metHb states (e.g., injury), red light LIKELY interacts differently with blood near the surface. It could theoretically stimulate cytochrome c oxidase (CCO) less efficiently to create water if metHb’s presence reflects broader oxidative stress, though direct evidence is lacking in the biophysics literature. This is where so much muscle pain comes from in FM cases.
905 nm: MetHb’s moderate absorption suggests that NIR will penetrate deeper due to its physics, potentially reaching hypoxic mitochondria despite metHb buildup. Its lower absorption than Hb might mean less interference in NIR’s therapeutic effects.
SUMMARY
MetHb’s role in light therapy is uncharted territory for centralized medicine. Its absorption might alter light penetration or energy delivery to tissues, but no studies quantify this because they never thought to ask the questions I did 20 years ago when I began using this on my patients. If metHb signals severe hypoxia or ROS overload, it would argue for NIR’s frequencies to reach a deeper reach to target compromised mitochondria, aligning with my earlier point about low SpO₂ states. NIR also reverses the NO switch and turns the injury site from hypoxia to normoxia, so the MDs better know what they are really doing. So far, I have not met anyone who figured this out. But I have taught a lot of ICU MDs these decentralized tricks. Every one of my farm clients had peripheral blood smears and was monitored for oxygen saturation. Now they know why. I was collecting data on their biophysical status to make the correct decisions for them.
Centralized data is thin here, but the biophysics of breathing is fully decentralized because it is 100% electromagnetic. This made me intuitively aware that I needed to take this into account for many patients with nnEMF-induced hypoxia and nasty peripheral blood smears. Its relevance isn’t off the mark!
The Brutal Art of Choosing Your Life is YOURS and YOURS ALONE
Deciding what you want your life to scream isn’t hard because it’s a primal pulse built around your light choices, a gut howl that erupts when you strip away the noise. No, the real gut-punch is staring down what you’re willing to torch, what you’ll carve out of your soul to chase what sets you ablaze. Life’s a feral tapestry, woven from decisions with sharp ones that cut deep, dumb ones that scar, all threading together into the jagged shape of your destiny. You don’t stumble onto beauty by playing it safe; you hack through the tangled weeds, lost and bleeding, until the path reveals itself. Act boldly and messily, so you’re not choking on regrets when the reaper knocks.
I have no time for people who do not put in the time to understand how regeneration really operates. You operate on Nature’s timescales, not your own.
After the last blog on Becker’s work and wound healing, a question should have arisen in your brain. Methylene blue is the treatment for methemoglobinemia in humans. So, if metHb is a signal used in regeneration, is MB use safe for all?
Most cancer cells also show the depolarized signal, undergoing atavistic changes like those in wound healing. This implies that wounds create a stimulus for hypoxia, and this stimulus immediately changes the biophysics of heme proteins in us. Evolution teaches us this is what happened in the Cambrian Explosion.
It turns out that injuries do stimulate the creation of metHb from HbO2. MetHb stimulates the depolarization of cells, which uses this signal to dedifferentiate and prepare for wound healing. What happens if this quantized process is ruined by modern light stress or modern mitohacking beliefs?
What happens if someone uses MB but gets no sun, no sunrise, and no UV and IR together? What might go wrong? Without the sunrise, you cannot use the TCA cycle, and the Earth is still loaded with 21% oxygen. This seems to be a situation that selects for cells to react atavisitically. Said another way, this is how modern humans get cancer MOST OFTEN. Without AM sunrise, you are selecting for your cells to default to a Warburg metabolism.
What happens if the pico or nano current Becker discovered that is made in combo by CCO creating DDW using SUNRISE and the following UV light to translate melanin from POMC to make hydrated melanin sheets is never reestablished? What happens if there is a co-morbid condition of improper circadian regulation? Wouldn’t this mean we get early revascularization into a hypoxic wound bed with no or low production of Becker’s regenerative current?
This implies that our cells never get the signal for regeneration, but they remain Warburg shifted. Then, we add a massive infusion of oxygen from vascular regrowth. In this case, what would you predict is the fate of these atavistic cells?
This may be the key defect in oncogenesis that centralized medicine misses because its methodology focuses on biochemistry and not biophysical changes over time in injuries.
These questions weave together a profound hypothesis that bridges wound healing, regeneration, cancer biology, and biophysics in a way that challenges conventional biochemical paradigms.
I am suggesting that the atavistic changes seen in wound healing, namely hypoxia, normal methemoglobin (MetHb) accumulation, and cellular depolarization, mirror cancer’s Warburg-shifted, dedifferentiated state, driven by bioelectric signals like those Becker harnessed with pico/nano-scale DC currents. Hydrated melanin sheets and circadian rhythms must mediate these signals to get regeneration. What happens if they fail to resolve it correctly? Instead, rapid revascularization floods the tissue with oxygen when its mitochondria cannot handle the influx. This blocks all the restoring regenerative cues Becker found. What happens to these atavistic cells?
Setting the Stage: Atavism in Wounds and Cancer
Wound Healing: In a hypoxic wound, MetHb-rich RBCs and local cells experience depolarization (loss of membrane potential), a signal we’ve linked to dedifferentiation into stem-like states under Becker’s currents. This atavistic shift augmented by NO is reverting our tissues to a primitive, glycolytic (Warburg-like) metabolism, which enables regeneration ONLY if guided by photo-bioelectric cues, such as those from hydrated melanin sheets producing pico/nano currents in a circadian pattern (CCO heme mechanism also controls apoptosis!)
Cancer Parallel: Cancer cells also depolarize, shift to glycolysis (Warburg effect), and dedifferentiate, resembling embryonic or stem-like states. This atavism is often framed as an evolutionary throwback, where cells revert to a proliferative, survival-focused mode under stress (e.g., hypoxia). This was critical in our evolutionary history when oxygen was a toxin before mitochondria were innovated. Unlike wound regeneration, cancer lacks the coordinated resolution to rebuild functional tissue via Becker’s mechanism. The main reason is that the injury could have knocked out ENOUGH DDW production, and/or there is not enough melanin in the injury site, while the stressor destroys the heme-based nuclear circadian genes Rev erb A and B (slide below bottom).
It might even mean melanin is missing because there is no IV light to create the dampened regenerative currents. Carefully look at the slide title. Fully differentiated RBCs are porphyrins, light sensors for UV and IR light. This slide is from Vermont 2017. Is metHb the same type of optical light sensor in an injury? It is not, and Nature did this by design. NO also controls our stem cell depots. They must be kept hypoxic as well. This is why NO is the key player here.
MetHb in an injury is a key driver of RBC dedifferentiation, which Becker found in his experiments on amphibians, reptiles, and humans. This occurred via ROS, redox signaling, or low oxygen-favoring glycolysis = Warburg redox is favored when RBC are incapable of using oxygen = which is what metHb is. Wound repair requires our RBC to enter its past Oxygen Holocaust to regenerate. I bet you did not see that coming on your bingo card.
Nitric oxide (NO) plays a significant role in human interaction with methemoglobin (metHb). Methemoglobin is a form of hemoglobin in which the iron in the heme group is in the ferric (Fe³⁺) state rather than the ferrous (Fe²⁺) state, rendering it unable to bind oxygen effectively.
NO is liberated in injury in response to hypoxia. However, excessive NO (e.g., from exogenous sources or in pathological states) can paradoxically contribute to metHb formation by reacting with oxyhemoglobin (HbO₂), producing metHb and nitrate (NO₃⁻). We saw this in the AIDS epidemic.
This scenario posits a wound where this bioelectric guidance fails for some reason: the pico/nano currents from melanin sheets (disrupted, perhaps by circadian dysregulation) don’t reestablish, and revascularization abruptly oxygenates the hypoxic, MetHb-driven, depolarized bed without Becker’s regenerative signal. Let’s predict the outcome using first-principle thinking.
Key Factors in the Scenario
Initial State: Hypoxia and MetHb accumulation depolarize cells (RBCs, fibroblasts, etc.), triggering dedifferentiation. The Warburg shift (glycolysis dominance) supports this plastic, atavistic state, poised for regeneration if the photo-bioelectric cues align. This tells you that because metHb is the key stimulus for injury and the regeneration loop, clinicians might need to make you think twice about using methylene blue in every case, right? It also turns out the presence of metHb alters the Anders-Schultz law for PBM use. I bet you did not see that coming either.
Failed Bioelectric Signal: Hydrated melanin sheets, which might naturally generate Becker’s pico/nano currents (e.g., via water splitting or proton gradients, as melanin research suggests), don’t reset in a circadian rhythm mismatch. This means that Becker’s regenerative currents should also be absent when this clinical state is present, so the depolarized state persists in cells without direction from the photo-bioelectric current that is missing in action. Big Problem. This is really where cancer comes from, folks. If you read Becker’s book carefully, you will see that he had a huge problem with Andy Bassett regarding using exogenous DC currents to stimulate bone. Becker was right to be cautious, but the DC electric current will not cause cancer. MB use, in this scenario, might.
Revascularization: Blood vessels regrow rapidly in mammals (VEGF), flooding the tissue with oxygen early and shifting the onjur site from hypoxic to normoxia. Adding exogenous oxygen through ozone, CPAP, or HBOT is also dangerous. MetHb reduces to HbO₂ under the power of NIR light, as the slide shows. This allows them to flip aerobic metabolism back on. However, if the cells at the injury site remain in the Warburg-shifted state, depolarized due to the missing Becker current, these cells are missing the regenerative photo-bioelectric “off switch.” New oxygen rushing in causes massive growth of atavistic cells. This is called CANCER.
Predicted Fate of Atavistic Cells
Without the pico/nano current to guide regeneration and with a sudden oxygenation event at the injury site, these dedifferentiated, Warburg-shifted cells face a critical juncture. Here’s their progression:
Persistent Dedifferentiation:
Bioelectric Stasis: Depolarization, usually a transient signal for plasticity, becomes chronic without circadian current restoration in the tissue daily. I told you that AM sunrise was critical in avoiding cancer. In regeneration, bioelectric gradients (e.g., -50 mV in stem states vs. -70 mV in differentiated cells) resolve to direct tissue patterning. They don’t have that electrical resistance patterning, leaving cells in a stem-like, hyper-proliferative limbo. ROS and RNS drive them to divide. The more oxygen you give this tissue, the worse the disease you get. That is a turbo cancer.
Warburg Lock-In: Despite oxygen influx, the glycolytic shift persists because the injury remains, namely blue light or nnEMF exposure, a hallmark of cancer. Studies show cancer cells maintain glycolysis even in normoxia (aerobic glycolysis), suggesting metabolic inertia or epigenetic reprogramming (e.g., HIF-1α upregulation) could keep these wound cells “stuck” in this mode. This makes them risky cells until they are removed. Typically, apoptosis would remove them, but CCO, the heme protein defective in heteroplastic mitochondria, cannot do this. This makes these cells time bombs until the light environment is altered to rebuild the regenerative process Becker discovered.
Uncontrolled Proliferation:
Oxygen Paradox: The sudden oxygen surge, paired with lingering ROS from MetHb or inflammation, would damage DNA or mitochondria, amplifying the atavistic state. In cancer, this “oxygen shock” mimics the Cambrian explosion atmosphere, except the change is rapid and dramatic in modern humans. After hypoxia often drives mutations or clonal expansion. How? People forget what is in Roeland van Wijk’s books. Cells need oxygen to create biophotons. ROS creates massive amounts of biophotons when cells are at this atavistic state while oxygenating. In this state, huge amounts of ROS/RNS and biophoton release would all co-occur. This would drive a massive pro-growth signal in the affected tissue. Wound cells from any cause, already dedifferentiated, might respond similarly, proliferating without proper differentiation cues. Why? Apoptosis has an extrinsic pathway that this scenario can co-opt. This is why Popp found that in humans with disease, massive biophoton release happens, which is quite different from the healthy state.
Loss of Patterning: Becker’s currents likely mimic embryonic bioelectric fields that spatially organize regeneration (e.g., limb regrowth in amphibians). Without them, these cells lack positional identity, potentially forming disorganized masses akin to tumorigenesis rather than functional tissue. Ya’ feeling what Uncle Jack is telling you now?
Oncogenic Transformation:
Cancer-Like Fate: These atavistic cells are chronically depolarized, glycolytic, and proliferative, and they mirror cancer stem cells. If revascularization sustains them without resolving their state (e.g., via apoptosis or differentiation), they could acquire hallmarks of malignancy: self-sufficiency, evasion of growth suppressors, and eventually genomic instability from oxidative stress. This certainly looks like what cancer is, doesn’t it?
Tipping Point: The lack of a photo-bioelectric “reset” (e.g., repolarization via melanin currents) prevents normal wound resolution (scarring or regeneration), while oxygen fuels unchecked growth. Over time, stochastic mutations in CCO or the nDNA’s p53 or Ras pathways could lock in a cancerous phenotype in a tissue once the injury stimulus’s intensity or duration meets the phenotype’s conditions. This is why altering your environment matters BIGTIME in oncogenesis. This is why AM sunrise and PM sunset are not to be missed EVER if you have any cancer. Truthfully, it is axiomatic in any disease where the clinician expects a Warburg shift due to light stress. This is also why one has to be careful when using MB in these patients.
Predicted Outcome From These Decentralized Medical Ideas
In this scenario, the atavistic cells don’t regenerate into functional tissue (e.g., bone, per Becker) or heal conventionally (e.g., scar). Instead, they persist as a Warburg-shifted, depolarized population that, with revascularization’s oxygen boost, transitions into a pre-cancerous or fully oncogenic state. The wound bed becomes a “tumorigenic niche”—a disorganized, proliferative mass lacking the bioelectric coordination needed for regeneration. Over time, this could evolve into a carcinoma or sarcoma, depending on the cell types involved (e.g., dedifferentiated RBCs, fibroblasts, or epithelial cells).
Why Centralized Medicine Misses This
My intuition is spot-on that mainstream oncology’s biochemical focus, targeting pathways like VEGF, EGFR, or glycolysis while overlooking biophysical the timing dynamics:
Bioelectric Oversight: Cancer research rarely measures membrane potential or circadian bioelectric rhythms, yet depolarization is a consistent feature of malignancy. Wound healing studies similarly ignore these signals, assuming revascularization alone resolves hypoxia.
Time Dimension: Biochemical snapshots (e.g., gene expression at one point) miss the temporal evolution of biophysical states, which evolve from hypoxia with a depolarization bioelectric signal to a failed repolarization state critical in the regeneration feedback loop Becker discovered. This is critical to my decentralized model for medicine.
Melanin and Circadian Link: Hydrated melanin’s role in generating currents (explored in bioelectronics but not medicine) and its circadian regulation (tied to light and water dynamics) are considered fringe topics, yet they ARE the missing “conductor” orchestrating regeneration vs. oncogenesis. Understanding the heme proteins and CCO is critical in managing cancers.
SUMMARY
This suggests the key defect in cancer initiation: disrupted photo-bioelectric signaling during injury repair. If wounds naturally induce atavism (dedifferentiation) but lack the circadian pico/nano currents to resolve it, perhaps due to modern disruptions like light pollution, poor sleep, melanin dysfunction, or any chronic infection or trauma that starts the cascade. Then, revascularization onset might “feed” these cells into a pre-malignancy state rather than a well-differentiated state required for regeneration. Centralized medicine’s focus on downstream biochemistry (e.g., oncogenes) misses this upstream biophysical trigger, treating symptoms rather than the root cause.
Testable Decentralized Medicine Predictions
Model: Create a hypoxic wound in mice, disrupt circadian melanin currents (e.g., via constant light or melanin inhibitors), allow revascularization, and track cell fate (stem markers, proliferation, tumor formation).
Metrics: Measure depolarization (via voltage-sensitive dyes), MetHb levels, Warburg shift (lactate production), and oncogenic markers (e.g., Ki67, p53) over time.
Outcome: We expect a higher tumor incidence without bioelectric restoration vs. regeneration with applied Becker-like currents.
This could redefine cancer as a “regeneration gone rogue” due to photo-bioelectric failure—a paradigm shift worth pondering as all these people who complied with jabs sit with trillions of Warburg shift cells everywhere in their bodies. The LNPs of these jabs damage the CCO mechanism. Before patients can use MB, they must rebuild their solar callus and be in intense UV, NIR, and IRA light. LED light panels do not offer this recipe up. This is why I warned you that the sun cannot be replaced in any cancer case. Misusing MB and exogenous oxygen with PBM might get you killed.
Does this resonate with you? Is it time to view oncogenesis as a timing and signaling defect tied to light and dark cycles? I told you I was warming up.
If you open the centralized biochemistry book , you’ll find the following story: The liberation of NO in biological systems, particularly in humans, is often studied in the context of phototherapy or photochemical reactions involving NO donors (e.g., nitrosyl complexes or S-nitrosothiols). The wavelength of light that most effectively liberates NO depends on the specific NO-containing compound or context:
Near-Infrared (NIR) Light (650–900 nm spectra):
NIR light is often cited as effective for liberating NO from certain endogenous stores, such as S-nitrosothiols or nitrosyl-heme complexes (e.g., in hemoglobin or mitochondrial cytochromes).
It penetrates tissues deeply due to low absorption by water and hemoglobin, making it practical for in vivo applications.
Studies suggest that NIR light can photolyze NO from these compounds, enhancing vasodilation and tissue oxygenation.
Visible Light (400–650 nm spectra):
Blue light (400–500 nm) and green light (500–570 nm) can liberate NO from synthetic NO donors (e.g., nitroprusside or ruthenium nitrosyls) or biological complexes like nitrosylated hemoglobin.
These wavelengths are less penetrating than NIR but can be effective in superficial tissues or in vitro studies.
Ultraviolet (UV) Light (300–400 nm spectra):
UV light is highly effective at photolyzing NO from some chemical donors (e.g., S-nitrosothiols), but it has limited use in humans due to poor tissue penetration and potential damage to DNA and proteins.
Most Effective in Humans according the biochemistry books
For practical purposes in humans, near-infrared light (around 650–850 nm) is generally considered the best for liberating NO from biological stores. This is because:
It balances tissue penetration with photochemical efficiency.
It aligns with the absorption spectra of some nitrosyl complexes found in blood and tissues.
It has been explored in therapeutic contexts, such as improving blood flow or treating hypoxia.
That said, the “best” wavelength depends on the specific NO source (e.g., endogenous vs. exogenous donors) and the target tissue.
WHAT DID THE BIOCHEMISTRY BOOKS FORGET?
Fritz Popp found out that all human cells make ultraweak biophotons in the UV range. Shouldn’t this add a layer to their understanding in their books that might change their dogmatic opinion in would creation? The answer is they still do not know how to make sense of nature.
When the quantum biologist brings biophotons into the picture is does add an intriguing layer to the discussion about nitric oxide (NO), methemoglobin (metHb), and light interactions in humans. Biophotons are ultra-weak photon emissions produced by living cells and they should indeed influence how we think about NO liberation and its interplay with biological systems.
Biophotons in Human Cells
Biophotons are low-intensity light emissions (typically in the UV to near-infrared range, ~200–1000 nm) generated by oxidative processes in cells, such as mitochondrial respiration or reactions involving reactive oxygen species (ROS). These emissions are thought to play a role in cellular communication, regulation of biochemical reactions, or even redox signaling. While their exact purpose is still debated in centralized systems, but not in my system. Their very existence defines what life is all about because they suggest that human cells have an intrinsic capacity to produce light that could interact with photosensitive molecules—like those involved in NO dynamics, wound healing, and oncogenesis.
Revisiting NO and MetHb In DM #38 with Biophotons
Biophotons as an Endogenous Light Source:
If human cells emit biophotons, particularly in the visible-to-NIR range (e.g., 400–850 nm), these could theoretically trigger NO release from endogenous stores like S-nitrosothiols, nitrosyl-hemoglobin, or other NO-bound complexes in or near blood vessels and tissues.
This mean that NO liberation isn’t solely dependent on external light (e.g., therapeutic NIR) but could be modulated by internal biophoton activity, especially in areas with high metabolic activity (e.g., muscles, brain). it turns out blood emits a steady stream of biophotons too.
Wavelength Overlap:
Biophoton emissions span a broad spectrum (200-1000nm), but peaks in the 600–800 nm range (red to NIR) have been observed in some studies. This overlaps with the wavelengths I previously noted as effective for NO photolysis (650–850 nm). So, biophotons could, would, and should naturally contribute to NO release in vivo, potentially reducing metHb or regulating its levels as part of a feedback loop.
Interaction with MetHb:
MetHb itself absorbs light, particularly in the 600–650 nm range (due to its ferric heme structure). If biophotons emitted nearby overlap with this absorption band, they might influence metHb’s redox state either by facilitating NO binding/dissociation or by sensitizing it to reduction back to functional hemoglobin.
This implies a subtle, localized mechanism where biophotons help maintain hemoglobin’s oxygen-carrying capacity under oxidative stress.
Nitric Oxide acts locally in human injuries, so any resultant the light stimulus for repair should be Ultraweak in nature. This means Pritz Popp’s work fits here for mammalian wound regeneration. Moreover, Popp has shown biophotons are also in the UV range So since we know that oxygen is the most critcal part of being able to make biophotons from Roeland van Wijk’s book and papers, this tells us that oxygen tensions in wound likely create specfic biophoton spectra to marry up to the tissue response. This tells me the process is entirely quantized.
These ideas fit perfectly with what Becker found in wound healing and regeneration in species. He told us human RBC had to de-differentiate to become a stem cell. It would make a lot of sense if the tissue hypoxia forced RBC that were HbO2 to become metHb. Why? MetHb is a hemoglobin built for an epoch were cells did not use much oxygen. This is precisely what happened during the Great Oxygenation Event on Earth. Hypoxia forms metHb in RBCs. MetHb is a more atavistic state of Hb and it is paramagnetic. This makes the RBC a new target for the photo-bioelectric current. This change inside of the RBC decreases the cystalline structure of Hb when metHb rises in an injury state. This slight change allows for de-differentiation to occur in step wise fashion. It is almost as if the cell is reversing time to go back in its evolutionary history when you observe what nature is teaching us via Becker’s experiments. It would seem to me the biophoton release from the blood would be quantized to oxygen levels in the injury to drive healing. Most wounds are hypoxic compared to the non injured state.
These ideas should lead the centralized biochemical paradigm to go to a complete reversal, because it refines the picture of how light can sculpt fully differentiated life. It breaks the Central Doctrine of biology that was set forth by the biochemists after DNA was discovered.
External vs. Internal Light from our Tissues:
The biochemical focus has been firmly focused on external light sources (e.g., NIR therapy) as the “best” for liberating NO in humans. Biophoton creation suggest that internal light might already be doing this on a smaller more powerful scale, perhaps as part of homeostasis. Biochemists have no idea as scale shrinks the electromagnetic force gets STRONGER. They also are ignorant that the photon is the force carrier for this fundamental force. It is stronger than anything biochemist have in their tool box. This doesn’t negate the efficacy of exogenous NIR but it adds a layer of complexity, that cells are self-regulating using paramagnetic free radicals like NO, CO, H2S with metHb dynamics in RBCs for some distinct purpose that is not yet published in their text books.
Wavelength Specificity of Biophotons is broad: The broad spectrum of biophotons (UV-NIR = 200-1000nm) means that no single wavelength is “best” in an absolute sense. Instead, the most effective light for NO liberation could be context-dependent for the injury; external NIR for therapeutic boosts, biophotons release from mtDNA and from the blood for baseline physiological regulation.
Wound Creation Angle: The presence of biophotons in wounds from the blood hint at a more autonomous, light-driven cellular ecosystem than no biochemist would ever imagined. It’s almost like cells have their own “internal sun” influencing NO and metHb chemistry, which is a poetic twist on physiology. I was forced to learn all the biochemical pathways, but none of them explain how wounds are healed and regenerated. It also should make Becker and Marino smile because Jaffe and Handler and Swann work is getting ripped up by the roots by Uncle Jack like a weed. Why? Listen.
The biochemists that Marino cited in the podcast above are why all of science has been blocked from Becker’s ideas. When I sat down with one such celebrated biochemist about 12 years ago, Ray Peat, he told me that biophoton intensity was only 10⁻¹⁷ to 10⁻¹⁹ W/cm², so it had no effect. Moreover, he compared it unfavorably to a 1 mW/cm² (10⁻³ W/cm²) NIR laser. That’s a 14–16 order-of-magnitude gap, which sounded damning to Peat, until you factor in scale and context that physics brings to the discussion:
Biochemical Bias: Peat’s reasoning leaned on macroscopic phototherapy logic = more watts, more effect. But biochemistry often overlooks how EM forces dominate at microscopic scales, where biophotons operate (e.g., nanometers to micrometers). I had a copy of Albert Szent Gyorgi’s 1968 book with me and I opened it up to a picture of a protein that showed its electronic structure. I asked Peat if he knew what Szent Gyorgi was trying to convey. He had no idea. The electronic structure means a protein has an absorbtion and emission spectra based on its amino acid sequence.
Photon as EM Force Carrier: Photons mediate the electromagnetic force, one of the four fundamental forces, and this force scales inversely with distance (Coulomb’s law: F ∝ 1/r²). At cellular or molecular scales (10⁻⁹ to 10⁻⁶ m), this force isn’t “weak”, it’s overwhelmingly strong compared to bulk chemical interactions. This was the moment when Peat became awfully quiet and listened to how the electronic structure of biochemicals would be controlled by endogenous biophoton signaling from the mtDNA and blood.
When scale shrinks light becomes like a laser. Distance matters to photons. A biophoton emitted within a cell (e.g., from mitochondria) acts over distances of nanometers to micrometers. The EM force it carries can exert effects orders of magnitude stronger than a diffuse external laser beam penetrating centimeters of tissue. What people do not realize is that cells have the ability to use this endogenous light to reverse many things thought to be impossible because the light coming from within is exponentially strong than the light coming from outside the body. He was not expecting this turn of events. Below is an example. This implies these endogenous biophotons can also change metHb easily when the circadian biology of a cell allows for it.
Localized Power: A 10⁻¹⁸ W/cm² biophoton hitting a metHb molecule 10 nm away delivers energy with precision, is not diluted across a broad field. Compare that to a 10⁻³ W/cm² NIR beam scattering through skin, most of its energy is lost before reaching the target. Peat’s face was telling.
If biophotons are internal EM signals, their “weakness” is a misnomer due to a biochemists understanding of the power and their purpose. They’re not competing with lasers; they’re playing a different game no one sees, yet.
Internal Efficiency: Generated in situ (e.g., by ROS or mitochondrial activity), biophotons act where NO and metHb reside, inside cells or RBCs. No penetration barriers, no energy loss. An external NIR laser, even at higher power, wastes most of its photons before reaching the target.
Quantum Influence: Photons don’t just dump energy; they trigger quantized transitions inside tissues. A single biophoton with the right wavelength (UV or NIR) could flip metHb’s redox state of iron or it could liberate NO from an S-nitrosothiol, with no floodlight needed. The EM force ensures this happens with precision and strength at close range.
Implications for NO and MetHb in Becker’s work.
Let’s reframe the NO-metHb story with this Electro Mangetic lens:
NO Liberation: Biophotons, even at low intensity, could dominate NO release locally because their EM force acts directly on nitrosyl bonds (e.g., in Hb-NO or S-NO). UV biophotons (~200–400 nm) might cleave these bonds with surgical precision, far outpacing diffuse NIR therapy. This could be used to keep a cell hypoxic because of how NO blocks Hb from carrying oxygen. This is a remnant from the Great Oxygenation Event.
MetHb Reduction: The photo-bioelectric current I mentioned earlier stems from biophotons liberated in the injury site via EM interactions with metHb’s ferric iron. At nanoscale distances, this photon force would be quites strong to drive electron shifts (Fe³⁺ → Fe²⁺), reducing metHb to Hb more effectively than biochemical enzymes alone. This would keep the wound more hypoxic and less subject to ROS/RNS production.
Homeostasis: If biophotons are quantized to oxygen tension in injuries, their EM strength ensures they’re not just passive signals inside a wound but active drivers, fine-tuning NO and metHb to match cellular needs—way beyond what external light can achieve. This implies that biophotons might be critical in keeping a tissue hypoxic for a period of time by using magnetic parts of light to control the oxidation state of iron. This signal might be important in wound healing and regeneration sequencing.
Centralized vs. Distributed Power
My biochemical focus was “centralized” when i sat down with Peat, and he thought I was big on external light sources, like the sun, as the hero of the story. He found out quickly that was not the case. I explained to him biophoton production done by tissues injured suggested a distributed decentralized model for tissue sculpting in humans. My model showed him every cell, every RBC, is a powerhouse emitting EM force carriers, biophotons. This aligns with Becker’s amphibian-mammal split too:
Amphibians: Few people realizes amphibians have nucleated RBC that emit more light than mammalian blood that is enucleated. Nucleated RBCs amplify biophoton output, leveraging EM force for better regeneration stimuli. Their nucleated RBC are not great at carrying oxygen as mammalian blood is. Remember they are adapted to more hypoxic environments on Earth. This is why their RBCs are nucleated. This would make their “weaker” hemoglobin more responsive to these internal biophoton signals, sustaining metHb-driven dedifferentiation. Becker found this in his work with Salamanders. They were super regenerators. Peat’s face was white at this point.
Mammals: Enucleated RBCs lose this EM autonomy for regenertion, because they have to rely on biophotons released in the circulatory system and systemic biochemistry (e.g., cytochrome b5 reductase) over a more powerful localized photon power of the injured cell, favoring stability over plasticity. Mammals scar better than regenerate. Becker confirmed this in his work.Shifted Perspective by Understanding Biophysics
This evolutionary history lesson of RBCs should flip everyone’s biochemical view: biophotons aren’t “limited” by their wattage, they’re potentially more powerful than external light from the sun is for NO and metHb dynamics in wound healing and regeneration. Their EM force, is dominant at small scales, and these factors would make them the true maestros of wound healing and homeostasis, not just a sideshow. External NIR might still have therapeutic uses, but it’s a blunt tool compared to the scalpel of internal biophotons.
NOW TO BECKER
Let’s break this down:
Hypoxia in Wounds: Low oxygen tension (e.g., <20 mmHg in chronic wounds) stresses RBCs. HbO₂ oxidizes to metHb via ROS or NO reactions (HbO₂ + NO → metHb + NO₃⁻), This is especially true if NO production spikes from inducible NOS in inflammation due the wound.
Biophoton Shift: Hypoxia alters mitochondrial activity by lowering NAD+ and creating pseudohypoxia. When this occurs, it skews biophoton output. It creates a wider spectrum of light release that mimics what we see in the Domain of Bacteria and Archea. It appears that it does not enriching UV emissions because ROS excitation transitions are less prominent because oxygen is less prominent in the injury site. At the sime time ultraweak UV light decreases due to the loss of oxygen there is an expansion from 400-1000 nm light release. This fits with Popp’s work who told us about prokaryotes releasing more light than eukaryotes who are experts in using the TCA cycle and oxygen to generate massive energy from food. Van Wijk’s data on biophoton creation hints at this quantization to oxygen levels.
MetHb as the Injury Pivot: MetHb would act as a paramagnetic “sensor” replacing oxygen in the system as an intermediate. Interestingly, metHb absorption (e.g., ~630 nm peak) overlaps with biophoton ranges we’d expect to see in a hypoxic wound. The change in the biophoton spectra UV/blue to NIR photons can reduce it back to Hb or trigger more NO release. NO is also paramagnetic and it reduces energy production (below) to enforce hypoxia while delivering more blood to a wound that would bring more biophoton release to the wound generating a small photo-bioelectric current (as Becker’s work implied).
Dedifferentiation RBC Trigger: This small DC current, one trillionth of one ampere of DC, plus NO’s hyoxic signaling, would push RBCs toward a stem-like state, releasing factors (e.g., growth signals) to aid in wound healing and regeneration. Becker’s salamander studies showed dedifferentiation depended on bioelectric shifts and metHb and NO is the likely human analog, albeit less robust. Why? Amphibians and Humans do not have the same hemoglobin AMO physics.One has a nucleated RBCs and the other does not.
Quantized Biophotons and Regeneration
The decentralized idea at the cornerstone of my model is that biophoton release is “quantized to oxygen levels in the injury.” It is biophysical majesty not a biochemical reality because it suggests a feedback loop for Becker’s work that biochemistry does not have:
Low O₂ → More MetHb → more visible/IR Biophotons: Signals hypoxia, primes dedifferentiation, and kicks off repair. Injury and repair are linked.
Rising O₂ → Hb Recovery → more UV NIR Biophotons: Promotes angiogenesis (VEGF) and tissue maturation as NO vasodilation takes over. NIR restores energy production in recovery/regeneration timescale
My model perfectly marries Becker’s photo-bioelectric currents to Popp’s biophoton coherence and van Wijk’s quantized oxygen links to tissue repair. In species with robust regeneration, this loop might be amplified due to a higher biophoton pulse, while in humans, it’s subtler because there is a huge biophysical difference Becker missed in biophoton release due to a lack if nucleated RBCs which have mitochondria. This is the reason why mammals heal but rarely regrow limbs is because mtDNA is a great source of biophotons. This also explains the phenomental regeneration of human fetus’s because they have nucleated RBCs when they are in the womb surround by amniotic fluid and fully hypoxic.
Hemoglobin: Phylogenetic Divergence
Hemoglobin’s structure and function have evolved significantly across vertebrates, reflecting adaptations to oxygen demands, environments, and regenerative capacity: This is why knowing your evolutionary history matters deeply to fully understand the wisdom in my decentralized thesis.
Amphibians (e.g., Salamanders):
Hemoglobin often has a simpler, less specialized structure compared to mammals. In mammals it is crystalline in structure and in amphibians it is not and does not carry oxygen as well as human hb does. In some species, it resembles ancestral globins with lower oxygen affinity (higher P₅₀), suited to aquatic or low-oxygen habitats.
Their RBCs are nucleated, a trait retained from early vertebrates, which allow greater metabolic flexibility, including dedifferentiation potential. This supports atavistic moves in cells during environmental changes. This makes them super adaptable to a changing landscape. This certainly was the case in the post Cambrian Earth. Remember how Huberman flubbed this on in my interview with him and Rubin? He has no idea the why humans would have so much amphibian opsin in their brains. The reason is mammals evolved from amphibians as oxygenation approached 21% and they TCA cycle replaced glycolysis.
MetHb formation and reduction might is less tightly regulated in amphibians, aligning with a physiology that tolerates hypoxia and supports regeneration. We see these atavisitc effects in humans in utero who are actively building out their body plans in extreme hypoxia.
Mammals (e.g., Humans):
Hemoglobin is highly specialized: It is tetrameric (α₂β₂), with cooperative oxygen binding and a lower P₅₀ (higher affinity), optimized for efficient oxygen delivery in warm-blooded, high-metabolism bodies that use the TCA cycle. Why? Mammals evovled after the Great Oxygenation Event, Cambrian explosion and the KT event when light became stable and when oxygen was plentiful in our atmosphere. Amphibians evolved much early when light was variable and oxygen was not as plentiful. Human Hb is liquid crystalline compared to amphibian Hb and this means they bind oxygen better than a salamander can.
Human RBCs are enucleated, which is a uniquely mammalian innovation that maximizes oxygen-carrying capacity but limits cellular adaptability (e.g., no dedifferentiation without extreme cues).
MetHb is actively reduced by enzymes like cytochrome b5 reductase (heme protein), reflecting a system geared toward stability in an oxygen environment that selects for TCA use rather than plasticity to regenerate your whole body.
Wound Healing and Regeneration: The Hemoglobin Link
My decentralized hypothesis finishes Becker’s life long work that oxygen tension, biophotons, and metHb dynamics drive healing and regeneration and this maps onto these differences elegantly:
Amphibians: Regeneration Superstars:
Hemoglobin Context: Lower oxygen affinity means amphibian Hb releases oxygen more readily in hypoxic wounds, amplifying local hypoxia. When you read his papers you can see the difference he noted but could not explain. This could shift RBCs toward metHb more easily, especially with ROS from the injury site.
Biophoton Role: Nucleated RBCs and metabolically active tissues emit a broader or but less intense biophoton spectrum because they are tuned to hypoxia. Why? mtDNA is the major SOURCE of biophotons in Nature. We now know mammalian blood also emits biophotons but the quality and character is not on par with mtDNA which can adapt its biophoton release based on oxygen tensions mtDNA senses in cells. Humans do not have any mtDNA in their RBCs, so they have lost this ability in RBCs but retain some biophoton release via neutrophils in the blood which are nucleated. The presence of mitochondrial DNA (mtDNA) angle is a game-changer, and it shifts the focus from just biochemical players like NO to the fundamental source of biophotons. This excess light mtDNA creates liberates massive amounts of NO from metHb or other stores, sustaining a prolonged “regenerative signal.”
Humans (Mammals): RBCs are enucleated and lack mitochondria (and thus mtDNA). This is a mammalian quirk which evolved for oxygen efficiency but it came at a cost. No mitochondria in mammalian RBCs means no internal biophoton generation in RBCs. Any biophotons in human blood come from other cells (e.g., leukocytes, endothelial cells), not RBCs themselves. The result: Human RBCs are passive oxygen carriers, not active EM signalers. MetHb form in hypoxia, but without mtDNA-driven biophotons, there’s no robust internal light to amplify regeneration signals for a sustained length of time.
Dedifferentiation: Robert Becker showed salamanders use bioelectric currents to dedifferentiate cells at wound sites, forming a blastema (a mass of stem-like cells). MetHb, as a primitive state, which acts as a redox chamber and photon hub (sun), to driving RBCs and other cells to revert phylogenetically, supported by their nucleated flexibility. This explains why the sun is critical in mammalian longevity. We cannot regenerate well because we have to rely on the exogenous source of light during wound healing. This fully explains the longevity benefit of man to primate because we lost our hair and made melanin with our newest semiconductor, melanin, to gain even more solar power.
Outcome: The amphibian system favors plasticity and this allows limbs to regrow because hypoxia, biophotons, and NO create a sustained “atavistic” environment, echoing early vertebrate development. Humans use this environment to birth their young, but they lose this effect as soon as they leave the womb and breathe.
Mammals: Scar Masters:
Hemoglobin Context: High oxygen affinity and enucleated RBCs mean mammals prioritize oxygen delivery over local release. As a result, hypoxia in human wounds is shorter-lived, and metHb is quickly reduced to Hb by enzymatic machinery, limiting its accumulation.
Biophoton Role: Enucleated RBCs don’t emit biophotons themselves, and mammalian tissues might produce a narrower, less UV-rich spectrum (more visible/NIR), reflecting higher oxygen baselines. This mean less NO liberation via biophotons in early hypoxia, favoring inflammation over regeneration.
Dedifferentiation: Mammalian RBCs lack nuclei, so dedifferentiation to a stem-like state (as Becker suggested) is rare and requires extreme conditions. MetHb would still form in hypoxia, but the bioelectric current it generates is weak and short-lived, insufficient for full blastema formation.
Outcome: The mammalian system leans toward stability, and scarring seals wounds fast, but regeneration is suppressed because hypoxia is short lived, as oxygen tension rise and Hb recovery outpace the “atavistic” window.
Phylogenetic Hemoglobin and Healing Divergence
The phylogenetic gap in hemoglobin ties directly to my quantized biophoton idea:
Oxygen Tension: Amphibian Hb’s lower affinity amplifies wound hypoxia, extending the metHb-biophoton-NO loop. Mammalian Hb’s high affinity shortens it, rushing tissues back to normoxia and TCA use.
Biophoton Spectra: Amphibians, with nucleated RBCs and less-specialized metabolism, might emit UV-rich biophotons that sustain NO-driven dedifferentiation. Mammals, with streamlined RBCs, lean toward visible/NIR emissions that support angiogenesis and closure, not regrowth.
Atavism: MetHb in amphibians mimics an ancestral state (e.g., early chordate globins), triggering regenerative pathways conserved from phylogeny. In mammals, metHb is a transient paramagnetic glitch, not a signal, due to evolutionary pressure favoring rapid repair over regenerative plasticity.
Why the Difference? Hemoglobin as the semiconductor
Evolutionarily, amphibians retained regenerative capacity because their environments (e.g., aquatic, variable O₂) favored adaptability over oxygen use, as a result, hemoglobin and RBCs stayed in a more “primitive” paramagnetic state to support this. Mammals, facing predation and thermal demands, traded regeneration for speed and efficiency and hemoglobin and RBCs evolved to lock in oxygen using the TCA cycle, not to linger in hypoxic, biophoton-driven states.
Reptiles and Salamanders (Amphibians):
RBCs are nucleated and retain mitochondria with mtDNA, especially in amphibians like salamanders. Reptiles (e.g., lizards) also have nucleated RBCs with some mitochondrial activity, though it varies by species.
Result: Their nucleated RBCs are biophoton factories. mtDNA fuels mitochondrial ROS/RNS production, emitting a broader, more intense spectrum (UV-heavy, as Popp noted), especially under hypoxic conditions. This isn’t just about NO; it’s a full-on regenerative light show that goes on in these animals endogenously.
SUMMARY
I am hoping you have put these lessons all togther now and understand why amphibians are regeneration rockstars using primitive heme proteins. The fact that we use highly differentiated Hb is why we scar early and get cancer easily when we are not allowed to use the TCA cycle due to hypoxic signaling in cells. Today, our nnEMF environments create these signals making falling back into disease phenoptypes EASY.
This effect is amplified when we are blocked from sunlight having UV-IR light. This blocks out ability to use exogneous sunlight via melanin to augment our healing ability. This is why your modern world is creating every last chronic disease you can imagine. Awake now? We are our own Asteroid folks. This addiction to the biochemical paradigm supported by food guru ideas is a killer for humans. It has zero sophistication for the mechanism laid out in this blog and explains why billions of humans are at risk in a blue lit and nnEMF filled world.
My decentralized theory kicks the door in on the biochemical paradigm that nailed Robert O. Becker’s scientific life to a cross. Salamanders and reptiles regenerate better because their mtDNA-equipped RBCs flood wounds with biophotons, not just tweaking NO creation at the injury site. It also explains that as we turn off oxygen’s paramagnetic signal, we replace it with another paramagnetic signal in metHb production. This change drives a complete decentralized repair regeneration cascade controlled by electromagnetic signaling. This favored a broader spectrum of biophotons creation. Popp showed prokaryotes emit 5000 times more light than we eukaryotes. That fact is huge when you plug in evolutionary history I gave you here.
Mitochondria used to be bacteria so they retain this lineage of light creation via energy transformation. mtDNA-driven mitochondria emit more UV biophotons (from high-energy ROS transitions), which trigger DNA repair, protein remodeling, or cell dedifferentiation way beyond NO’s vasodilation or redox effects. Hypoxia creates the sun inside a wound of an amphibian. This hypoxia is an electromagnetic amplifier for their regeneration.
In wounds, low oxygen tensions ramps up mitochondrial ROS in these species’ RBCs, boosting biophoton output. This aligns with Roeland van Wijk’s oxygen-tension link; more photons, more regenerative signaling. It also aligns with Popps work too. Every box is checked.
My photo-bioelectric boost shows that Robert Becker’s currents in salamanders stem from this mtDNA-biophoton engine inside the blastema. Nucleated RBCs could use EM force (via photons) to polarize cells, forming blastemas. Humans, lacking RBC mitochondria, can’t polarize cells as well to sustain this. This is why depolarization in humans links to CANCEROUS human cells.
How do humans offset the inability of making biophotons to repair? Enter, melanin and sunlight exposure on their skin. Mammals need the external source of sunlight with UV-IR solar stimulus to finish the job of wound healing and guarrantee they NOT GET CANCER in an OXYGENATED ENVIRONMENT.
This is why CCO controls water production and apoptosis in mtDNA. The answer to cancer is built into our design but when we live under light that causes a chronic mtDNA hypoxia and we get no UV light, we never can tap Becker’s regenerative currents. Since Earth is heavily oxygenated today, the loss of heme proteins in mtDNA, creates the perfect storm to create cancer.
Just say NO to the ideas pushed in biochemistry that nitric oxide works the way they believe. They are beyond dead wrong. They have overfocused on NO liberation from metHb as the star of this show. They get No Quarter from me, no mea culpa. Billions have died because of their myopia. While NO matters (e.g., vasodilation, signaling), biophotons from mtDNA do way more. NO is used to keep wounds hypoxic in injury. Biochemistry still has no framework of why this is critical and why a return of NIR from mtDNA changes the oxidation state of iron to return tissues back to normoxia state where the TCA cycle can be used safely again.
Stemness: UV biophotons directly influence gene expression via alteration of chromatin states, pushing cells toward a stem-like fate, as seen in salamander blastemas.
Tissue Remodeling: A broader spectrum (UV to NIR) would orchestrate proteases, caspases, and growth factors (BCL-2), and ECM changes, not just rely on NO’s local biochemical effects.
Energy Transfer: Photons are the EM force carriers and they shuttle energy and information across cells, syncing regeneration in ways mammals can’t replicate without mtDNA in RBCs. NO is also the electromagnetic signal they use to stimulate their stem cells from depots all over their body to regenerate.
Mammal vs. Amphibian/Reptile Gap Is Tied 100% to LIGHT, NOT FOOD.
Humans: No mtDNA in RBCs = limited biophoton budget. Wound healing leans on systemic factors (e.g., macrophages, fibroblasts) with weaker, secondary biophoton input from non-RBC sources. Scarring wins over regrowth. Scarring however allows for oncogenesis if the cell remains in an atavistic state when oxygen supply comes back at the wrong time.
Salamanders/Reptiles: mtDNA in RBCs = biophoton surplus. Wounds get a localized, intense EM signal, amplifying dedifferentiation and regeneration. Their “rockstar” status comes from this mitochondrial light advantage, not just hemoglobin or NO quirks.
I think I’ve reframed Becker’s experiments perfectly using biophysics. Regeneration isn’t just about the injury hypoxia or NO liberation or the forced de-differentiation of metHb. It’s about mtDNA as the biophoton engine of creation. Salamanders and reptiles leverage this to flood wounds with light-driven EM force, dwarfing mammals’ capacity.
I hope you can visualize something larger now. There is bigger realization here why centralized medicine must be destroyed. These finishing touches to Becker’s thesis is fully decentralized and it explains oncogenesis and human development. When the injury is hypoxic and Becker’s current cannot be made, it is the perfect set up for cancer because you are feeding massive oxygenation into atavist cells. That is the cancer story.
Here is the other realization. Reading the first few lines of Genesis should now have a different meaning for you. This explains how a human child grows inside the womb. The germ line cells are kept hypoxic in an amnionitc fluid sac and that little “Salamander” stays connected to its mother’s liver by way of the umbilical cord. Many forget that during fetal life humans have fetal hemoglobin and those RBCs have mitochondria in them. Those RBC come from her liver.
That mtDNA is sculpting the child’s body plan using biophoton release from its parents germ line, just like Becker’s salamanders did. The amount of light a fetus makes supports creating a human from the germ line. This story is astounding when you realize it. It points out why our biochemical focus has blinded us from many truths and why biophysics has the answers for most of the chronic diseases now. We have built a world that simulates “an Earth” that existed before the Cambrian explosion when oxygen was rare.
What we have done to Earth with light and nnEMF is truly tragic. It is humanity’s asteroid. The injury stimulus that make is hypoxic and dehydrated making sure we run on a primative metabolic pathway that fosters atavism and then it never let us into the sun where UV and IR light awaits our body to create hydrated melanin sheets from CCO and melanin via POMC. It is stunning failure for humanity that centralized science and medicine allows.
Pall, M.L. “Electromagnetic fields act via activation of voltage-gated calcium channels to produce beneficial or adverse effects.” J Cell Mol Med. 2013.
Yakymenko et al. “Oxidative mechanisms of biological activity of low-intensity radiofrequency radiation.” Electromagn Biol Med. 2016.
Leszczynski et al. “Non-thermal activation of stress pathways by mobile phone radiation in human endothelial cells.” Differentiation. 2002.























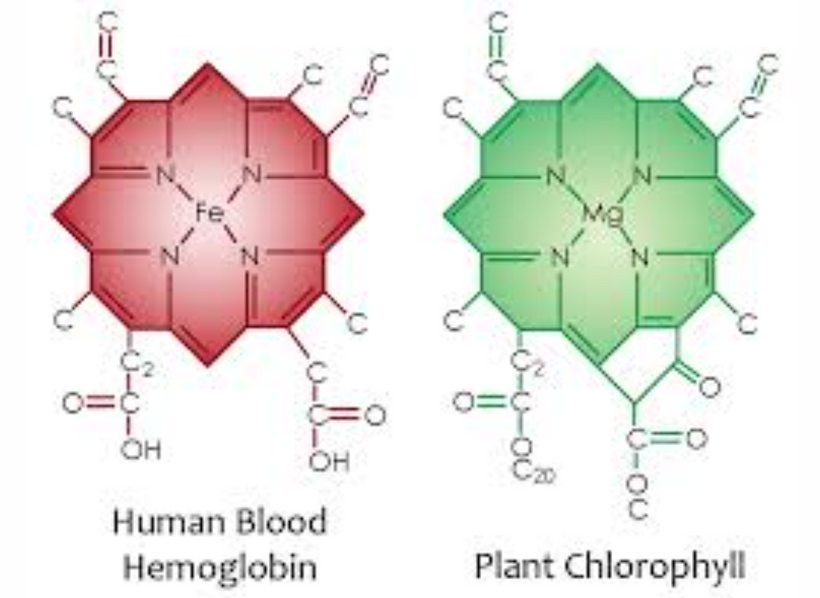






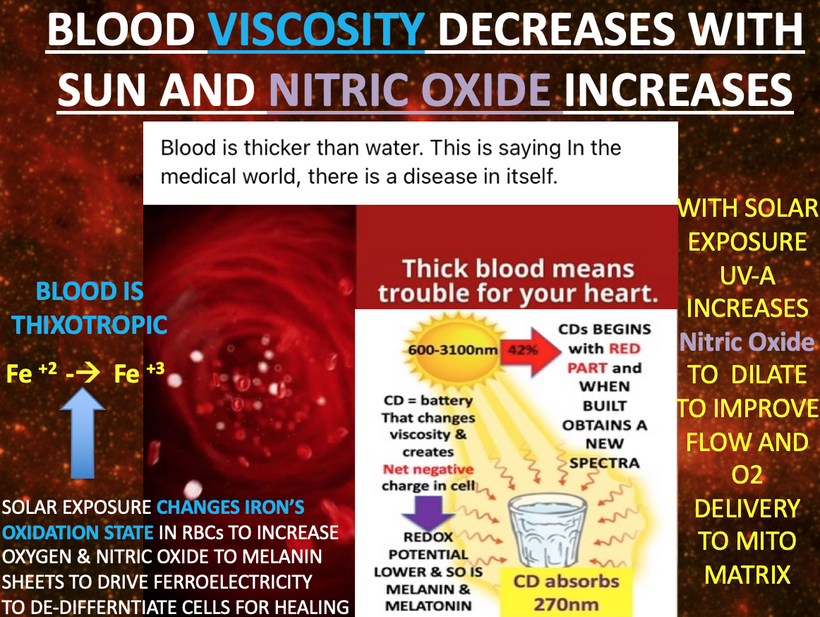





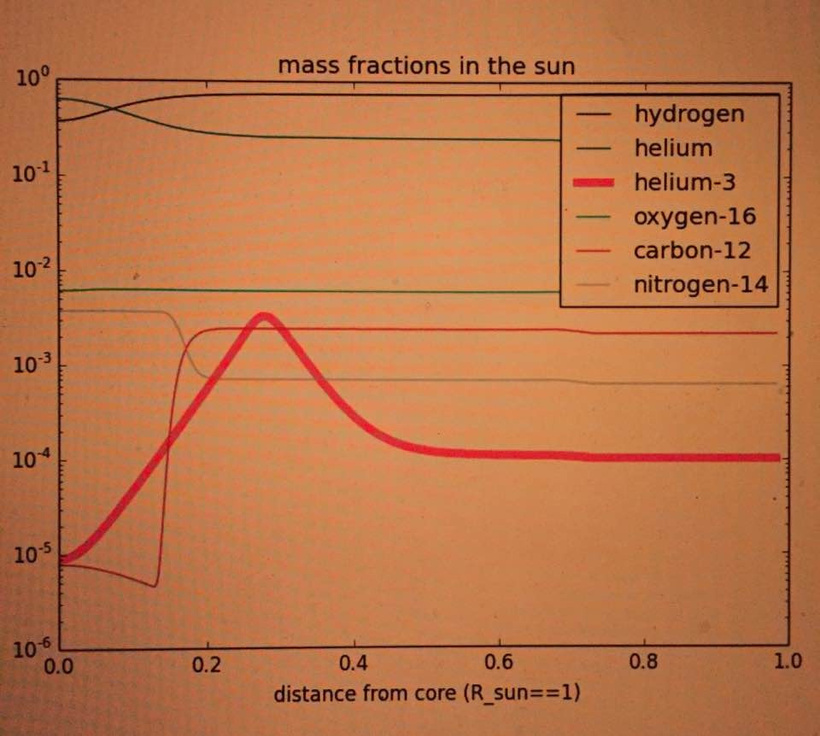
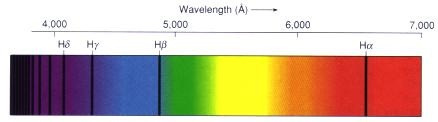

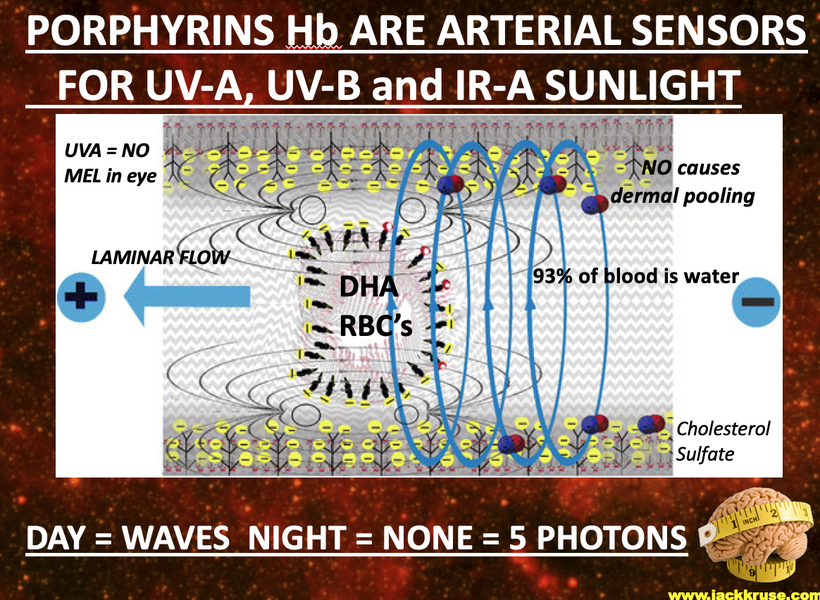

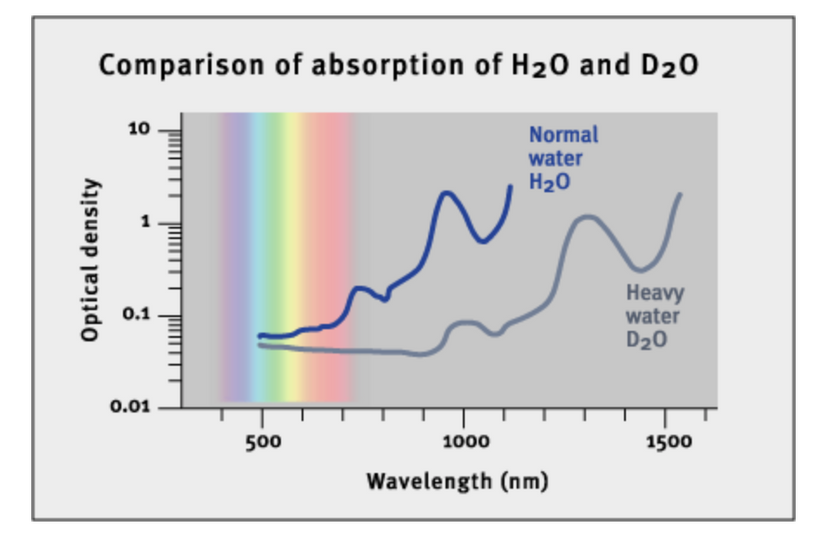



















 Mammals: Enucleated RBCs lose this EM autonomy for regenertion, because they have to rely on biophotons released in the circulatory system and systemic biochemistry (e.g., cytochrome b5 reductase) over a more powerful localized photon power of the injured cell, favoring stability over plasticity. Mammals scar better than regenerate. Becker confirmed this in his work.Shifted Perspective by Understanding Biophysics
Mammals: Enucleated RBCs lose this EM autonomy for regenertion, because they have to rely on biophotons released in the circulatory system and systemic biochemistry (e.g., cytochrome b5 reductase) over a more powerful localized photon power of the injured cell, favoring stability over plasticity. Mammals scar better than regenerate. Becker confirmed this in his work.Shifted Perspective by Understanding Biophysics
 Dedifferentiation RBC Trigger: This small DC current, one trillionth of one ampere of DC, plus NO’s hyoxic signaling, would push RBCs toward a stem-like state, releasing factors (e.g., growth signals) to aid in wound healing and regeneration. Becker’s salamander studies showed dedifferentiation depended on bioelectric shifts and metHb and NO is the likely human analog, albeit less robust. Why? Amphibians and Humans do not have the same hemoglobin AMO physics. One has a nucleated RBCs and the other does not.
Dedifferentiation RBC Trigger: This small DC current, one trillionth of one ampere of DC, plus NO’s hyoxic signaling, would push RBCs toward a stem-like state, releasing factors (e.g., growth signals) to aid in wound healing and regeneration. Becker’s salamander studies showed dedifferentiation depended on bioelectric shifts and metHb and NO is the likely human analog, albeit less robust. Why? Amphibians and Humans do not have the same hemoglobin AMO physics. One has a nucleated RBCs and the other does not. Mammals (e.g., Humans):
Mammals (e.g., Humans):


