CENTRALIZED VS. DECENTRALIZED MEDICINE
I visited a friend who was admitted to the ICU
The whole hospital is blue fuckin lit.
The wards were sealed tight, not a breath of fresh air.
The sun fights its way in, but thick glass blocks the full solar light spectrum.
WiFi routers & medical equipment blasting EMFs 24/7.
Every surface is sterilized, yet the air feels dead.
There are no negative ions or grounding; it is just recycled air that is heavy with chemicals.
Machines beep like a never-ending alarm.
Walls, lifeless and grey.
The only greenery?
Plastic plants collect dust in the corner.
Oxygen is pumped through machines, but there is not a single breath of fresh air.
The doors are locked as if nature is the enemy.
How the hell is this a healing place?
You go broke using the place.
It is a fiat hell hole to which everyone is sent.


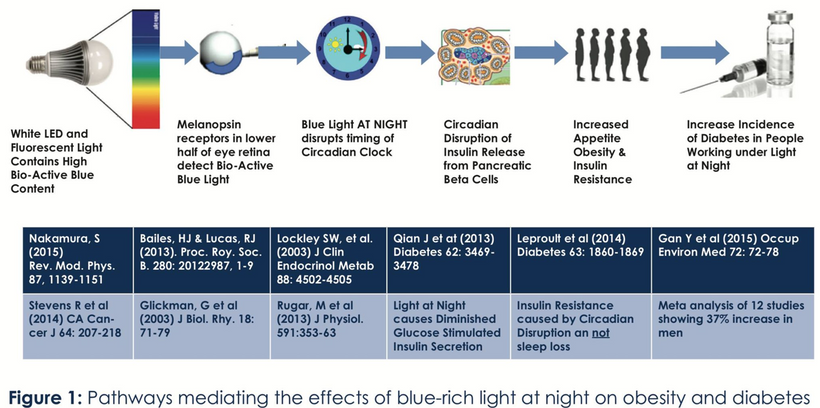
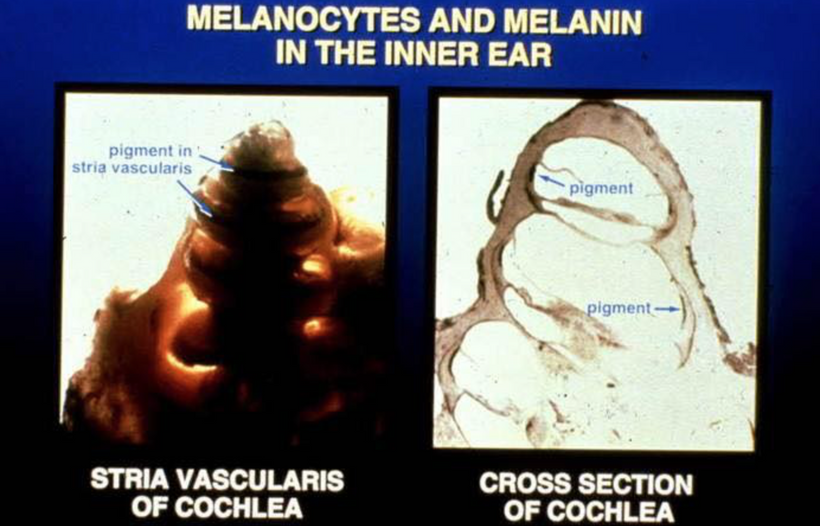
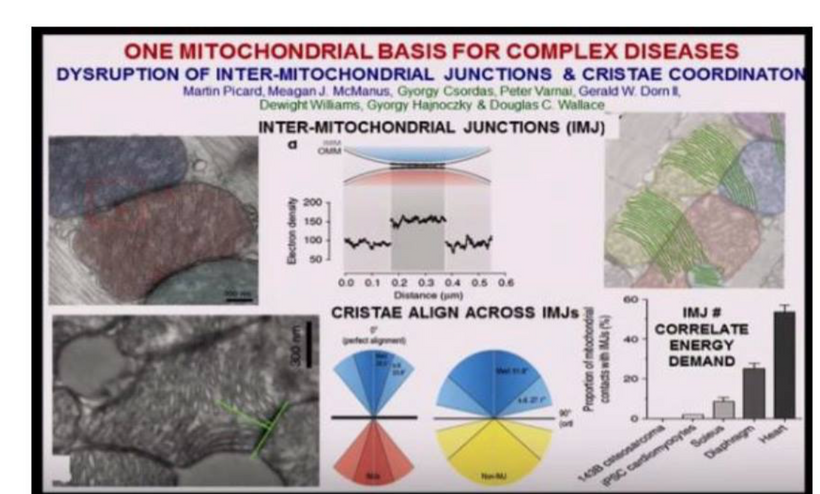
ALL CHRONIC DISEASES ARE LINKED TO THE WARBURG SHIFT
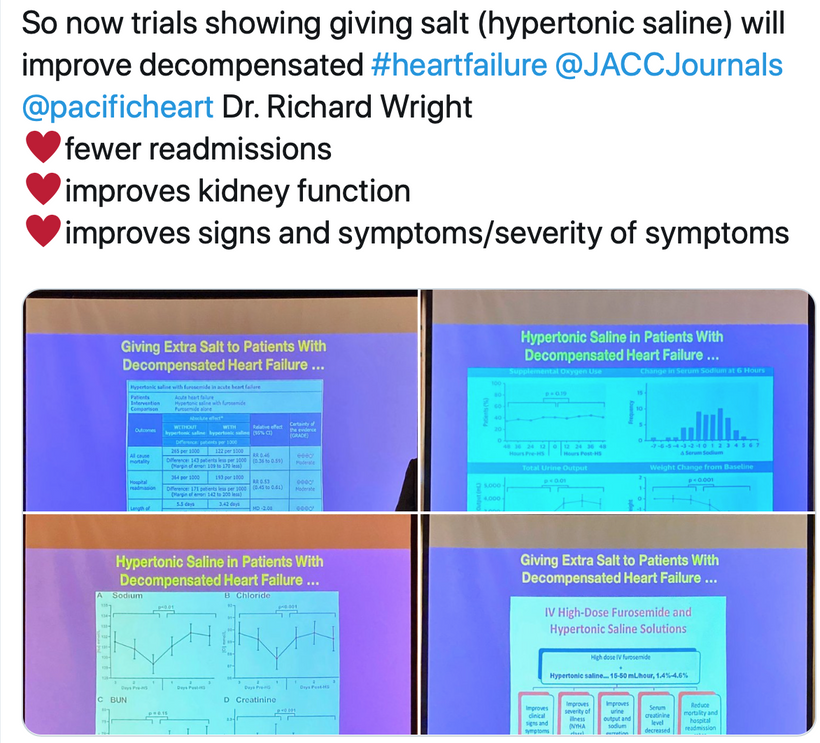
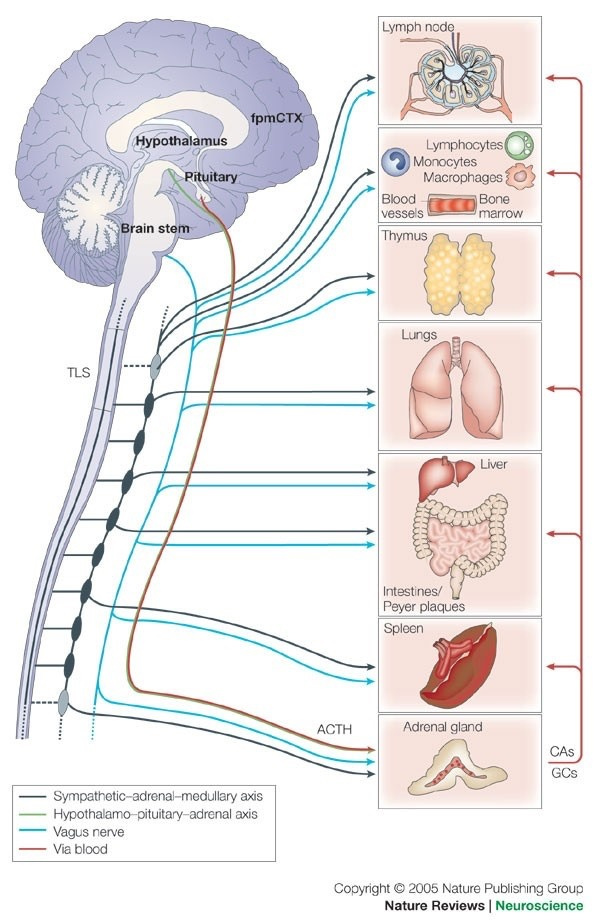
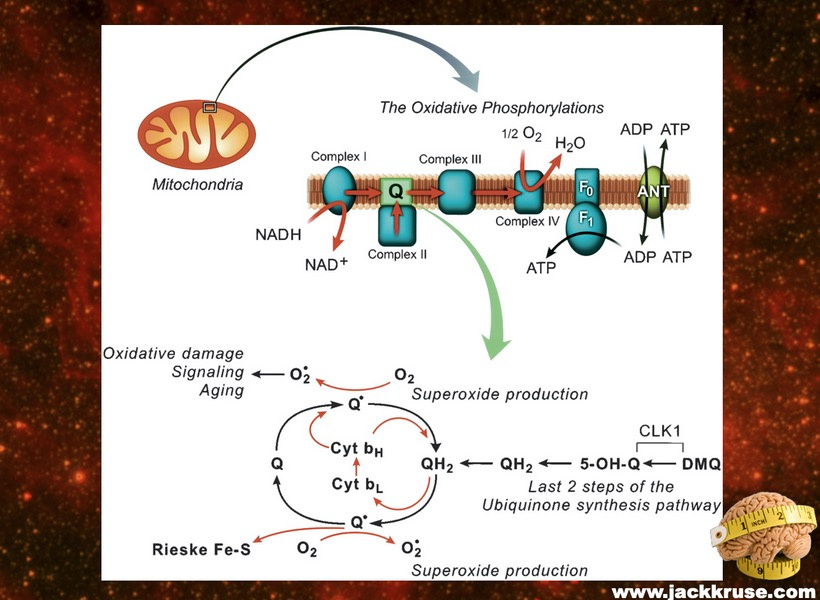
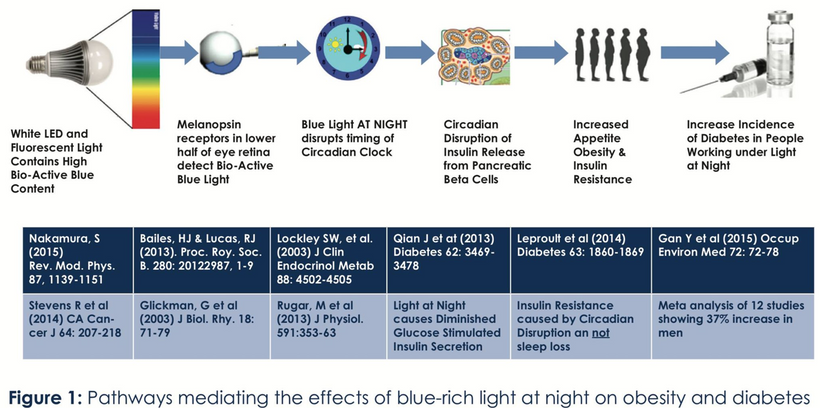
1. nnEMF-Induced pH Shift Drives Warburg Redox Shift: Light drives it, not food. The pictures above and below both show it in spades, but no one understands what these two pictures placed in blogs imply. The Warburg effect is characterized by increased blood glucose, glucose uptake, and lactate output, even under aerobic conditions. This reflects mitochondrial dysfunction (e.g., damaged IMM, cytochrome c oxidase inhibition). The damage of the IMM between NAD+ and oxygen allows for a 30 million charged field to escape into the cell. That charge then follows electric resistance pathways to cause many disease phenotypes. This is the stress response Selye talked about decades ago. He never was able to pin down how it began. All he knew was that it started on the HPA axis.
This podcast gave you the basics to the details in this blog: https://www.youtube.com/watch?v=IHSMqhrfvaU
The published science is precise that light stress by ALAN or a lack of sunlight drives metabolic shifts and how red light might serve as a therapeutic “drug” equivalent to counter the Warburg shift to change metabolism in ALL chronic diseases. In all stressors, vasopressin is the first stimulus to injury, and then the Orexin prism you learned about in QE #20 is next up in the cascade. Below, you see the definitive proof in two more pictures that red light reverses the effect of blue light exposure. This is what defines the Warburg Redox shift.

My photo-bioelectric hypothesis posits that nnEMF (non-native electromagnetic fields) and blue light damage melanopsin, mtDNA, and heme proteins, reducing DDW (deuterium-depleted water) production which leads to dehydrating melanin and increasing electrical resistance (éR) inside cells, leading to massive ROS/RNS rise, reductive stress (high NADH/NAD⁺), and Warburg metabolism (increased lactate). All of these cascades make oxygen a toxin because, due to its electronegativity, it pulls electrons across the IMM that is demolished. Why? With a stressor or trauma, excessive éR presents and leads to dissipative loss and bioenergetic inefficiency, reductive and oxidative stress, inflammation, molecular damage, and information loss. Thus, the hallmarks of disease and aging naturally arise from éR problems in the IMM on mtDNA. What you see and understand on the surface from biochemistry is not the entire biophysical story below built on the IMM.

Heme proteins are much older than mammals. This is a consequence of the evolution of our atmosphere. At the end of the Great Oxidation Event, heme regulation began to merge with POMC biology. Heme is a cofactor for many proteins, and it influences POMC expression via circadian and metabolic pathways. For example, heme binds to Rev-Erb-alpha and Rev-Erb-beta (nuclear circadian receptors), which regulate POMC transcription.
This links heme (and oxygen metabolism) to the light-driven circadian rhythms of mammals.

Heme proteins are much older than mammals. This is a consequence of the evolution of our atmosphere. At the end of the Great Oxidation Event, heme regulation began to merge with POMC biology. Heme is a cofactor for many proteins, and it influences POMC expression via circadian and metabolic pathways. For example, heme binds to Rev-Erb-alpha and Rev-Erb-beta (nuclear circadian receptors), which regulate POMC transcription.
This links heme (and oxygen metabolism) to the light-driven circadian rhythms of mammals.

THE DECENTRALIZED CASCADE TO THE ETIOLOGIES OF CHRONIC DISEASE
- Organisms as Open Thermodynamic Systems:
- Organisms are open thermodynamic systems dependent on energy flow (e.g., sunlight, food) and material exchange (e.g., waste export). Spent energy contributes to entropy, defining the flow of time. Molecular clocks (e.g., PER1/PER2) act as flowmeters of entropy, measuring cellular energy dynamics.
- Cells as Dissipative Structures:
- Cells escape the second law of thermodynamics (entropy increase) by creating order from chaos, functioning as dissipative structures. This order is built around the AMO (atomic, molecular, and optical) physics of atoms in cells, with molecular arrangement critical for function.
- Water’s Role in Cellular Organization:
- Water is critical in the molecular arrangement of biomolecules, influencing their physiological function. The movement of water (e.g., via hydration shells and gradients) changes biomolecular dynamics, mimicking semiconductor physics in silicon circuits (where electric power generates light). Cells use water to create and manage light (e.g., biophotons), maintaining atomic molecular order. Pure water contains the field of charge stored on the IMM between NAD+ and oxygen.
- Temperature affects semiconductors’ band gap size, too. You got this lesson in QE #29. Now you will find out how to link to your own disease.
- Cooling increases band gap size. Water changes the temperature of the semiconductive proteins inside of use, which generates electrical resistance.
- Cooling gave us evolutionary pressures to create catecholamine chemicals in prokaryotes like dopamine and adrenaline using a new semiconductive protein inside cells called melanin. When it degrades, it can become L-DOPA, which can become dopamine and adrenaline under a hypoxic stimulus.
- A lack of oxygen affected the early oxygen carrier semiconductive molecules called heme proteins. Recall that Robert Becker’s work around bone regeneration concerned itself with transforming RBCs to a pluripotential cell via a pico to nano ampere currents. The initial oxidation of hemoglobin to the ferric (Fe3+) state without oxygen converts hemoglobin into a useless “hemiglobin” or methemoglobin, which cannot bind oxygen.
- KEY POINT NO ONE REALIZES: This step exists in hemoglobin biology to deactivate hemoglobin’s ability to carry on its adult physiology to travel back in time atavistically to create Becker’s regenerative current. Becker’s de-differentiation experiment never looked at the blood cells that were being transformed. If you understood the evolutionary history of hemoglobin creation, this was a methodological error. It occurred during the GOE.
- Methemoglobin is an ancient protein used by life long ago when oxygen WAS NOT prominent in our atmosphere. This was when all life on Earth had an “allergy” to oxygen because oxygen was toxic to bacteria and archaea then. Methemoglobin naturally builds up in human injuries to precondition the cell to return in time. If the cell is filled with oxygenated hemoglobin, it cannot generate Beckers’ regenerative current. One thing that can be used to help this process is methylene blue. But MB has to be used at the right time. The first week is the incorrect time to use it. Methylene blue has two major effects that are beneficial for humans who cannot generate Becker’s healing currents.
- NUMBER 1: It alters the eR on the IMM in a damaged state, and
- NUMBER 2: it can increase NO delivery to tissues that become hypoxic with a lowered NAD+. This increase in NO allows mammals to access their stem cell depots to repair the damage in question. The stressor and damage are always linked to tissue-level hypoxia or pseudohypoxia, where NAD+ has dropped.

- Early versions of hemoglobins suffered from this problem = myoglobin. Later, mammals figured out a novel way to stabilize hemoglobin using green light when oxygen filled the atmosphere. First, I have to explain how we got there. Modern versions of hemoglobin in normal red blood cells are protected by a reduction system in the RBC by an aromatic amino acid (histidine) and a sea of electrons from the water in the blood. The surface of Earth was getting pounded by UVC light for long periods. Since life was prokaryotic and anaerobic 2.7 billion years ago when cyanobacteria evolved, it is believed that oxygen acted as a poison and wiped out much of anaerobic life, creating an extinction event of the old guard in life, including LUCA. LUCA = last unknown common ancestor.
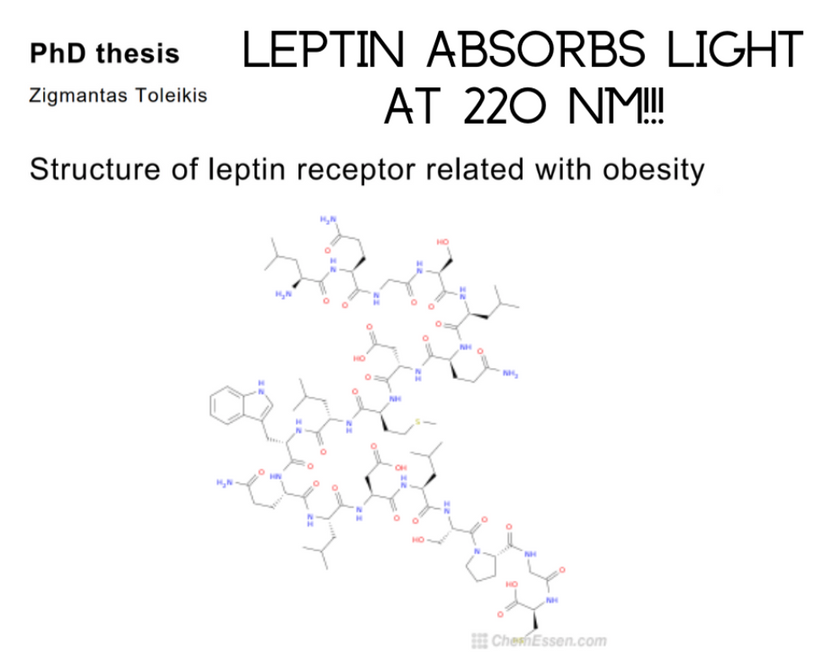
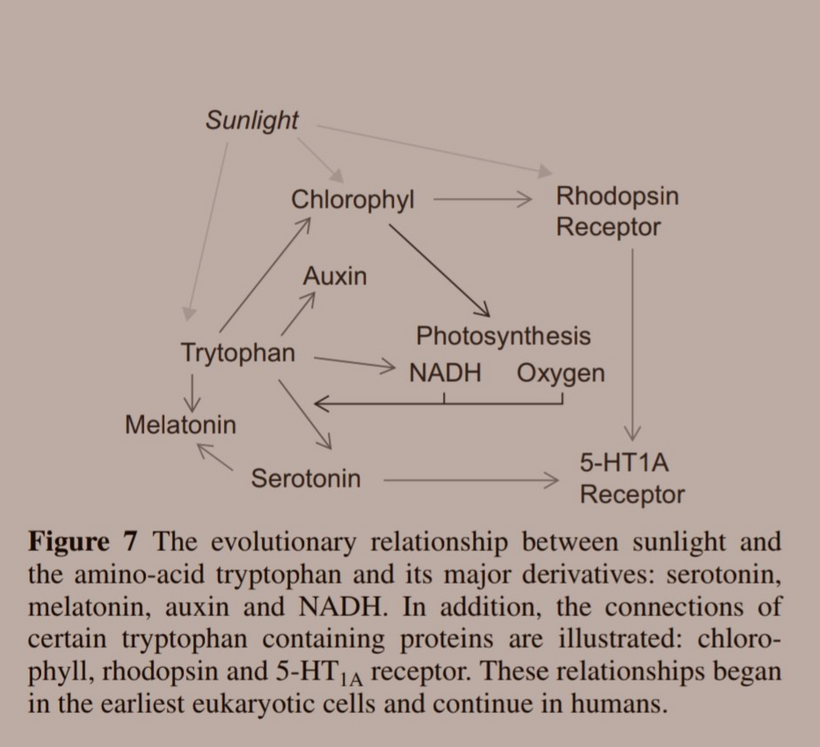 This environmental change drove evolutionary pressures to innovate proteins that used aromatic amino acids to build the most critical parts of the modern metabolism we see today in cells. Their absorption spectra can go from 150nm VUV to 400 nm UV-A light. Melatonin is one of the most ancient semiconductors known. Its functions have evolved as the atmosphere of Earth changed its atomic concentrations. This is why leptin has 220 nm absorption spectra when you look at it
This environmental change drove evolutionary pressures to innovate proteins that used aromatic amino acids to build the most critical parts of the modern metabolism we see today in cells. Their absorption spectra can go from 150nm VUV to 400 nm UV-A light. Melatonin is one of the most ancient semiconductors known. Its functions have evolved as the atmosphere of Earth changed its atomic concentrations. This is why leptin has 220 nm absorption spectra when you look at it
- Melatonin, NAD+/NADH, dopamine, adrenaline, leptin, epinephrine, etc. Light controls the flux of all these biomolecules because of the movements of H+ in cells. Remember, all enzymes that create these chemicals use proton tunneling to get the job done. Proton tunneling is linked to the HIF-1 alpha and PER2 gene that uses light to increase the periodicity of cell molecular clocks. Increasing periodicity = better clock management.
- Tryptophan, another aromatic amino acid, became very useful to cells living in an oxygenated world as a “time crystal” for cells because it has only one DNA codon, and its catabolism changes as light/dark signals change with the tilt of the Earth that gives us seasons. NAD+ and melatonin are both made from tryptophan. NAD+ and 95% of our melatonin is created in mtDNA. You might want to go back and re-read QT-14 now.

- Future life forms would need this information because oxygen gave us cool and hot seasons in one year as the Earth revolved. This cyclic pattern was built into cell metabolism as it got more complex, as the slides above show.
- Since oxygen has a high redox potential, it acts as an ideal terminal electron acceptor to generate energy after a nutrient breakdown. Oxygen soon became indispensable for metabolic activities. Organisms also evolved strategies to detoxify the reactive oxidative species that resulted from aerobic metabolism. Told ya’ that Vermont talk was important.
Though sequencing and phylogenetic analyses estimate the evolution of ROS-detoxifying enzymes even before the advent of aerobic microbes, the Great Oxidation Event acted as the catalyst to shape the directed evolution of enzymes like superoxide dismutase (above) and catalase. Catalase (above) is one of the earliest heme proteins, ancient myoglobin, and hemoglobins.
-
- Note the wide-band semiconductor components in the picture above: the Fe-S dopant semiconductors and their roles in creating the free radical signal in mitochondria. Below, note how mammals’ chromophore proteins are linked to aromatic amino acids, heme proteins, and melanin at some level. Can you guess why yet?
- As oxygen continued to mushroom, the high ionosphere became filled with a new gas that decreased the terrestrial solar spectrum. Life had to react to this, and it fueled changes in heme proteins that appeared on Earth’s surface in early life forms.
Oxygen was also responsible for forming the ozone layer in the atmosphere. The UV radiation from the sun split oxygen molecules (O2) into two atoms of oxygen, which then reacted with another oxygen molecule to generate ozone (O3). Ozone acts as a natural sunscreen for Earth to prevent harmful UVC and parts of the UV-B radiation from reaching the Earth’s surface. This reduction began the evolution of new semiconductors in the two DOMAINS of life on the surface of the earth called melanin.
As oxygen continued going higher, it fueled the Cambrian explosion, and life was able to take advantage of the mirror image of the photosynthetic arm of life on Earth. Namely, mitochondria developed. Complex life captured mitochondria in their tissues as a stowaway to transform solar energy into CO2 and water. It transferred electrons from food to oxygen to fuel this solar battery. At this point, life exploded, and all the complex life on Earth we know about today showed up almost overnight. Eukaryotes came from the fusion of the other two Domains in endosymbiosis. We believe chlorophyll and mitochondria were also innovated at this time.
Stress, Vasopressin, and Water Conservation:
- Stress and trauma (e.g., nnEMF, ALAN) disrupt water dynamics, triggering vasopressin release to conserve water. This mimics the VP-ISR-GDF15 axis activation in modern mammals. This is done because injuries all induce dehydration, and hypoxia of our semiconductors facilitates increasing éR, entropy, and cellular chaos, driving most chronic disease phenotypes. This mimics life during the GOE.
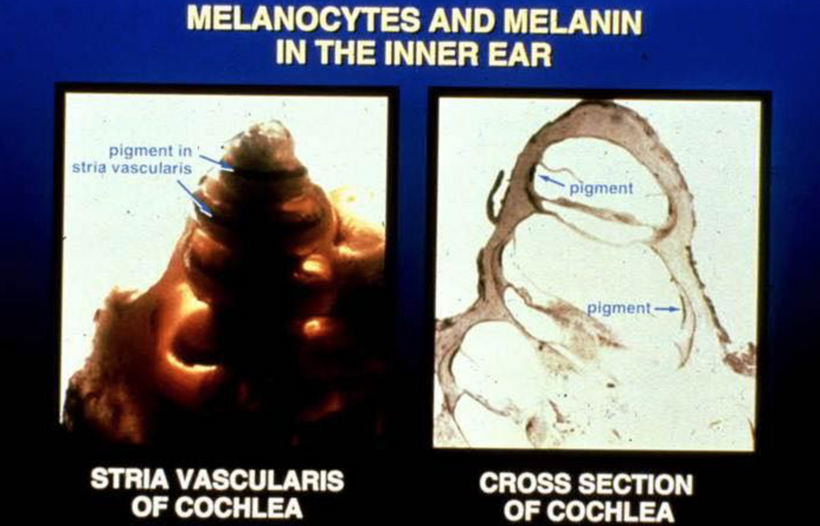
- When the human body is sick, diseased, and energy inefficient from any stressor or stimulus, its evolutionary directive is to eat itself removing all the diseased cells and organelles and cancerous cells. To do this, it needs to generate a regenerative DC to do so. This current relies on the ability of heme proteins to make water to hydrate melanin to create this small current. When the cell cannot do this for any reason, big problems await this organ. The disease begins to spread like an infection does through the organ. We call this heteroplasmy.
- HIF-1 and Cellular Hypoxia:
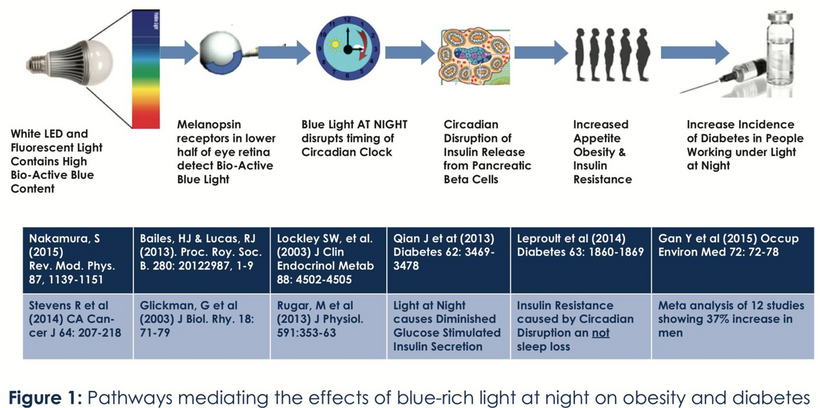
- Poor sunlight, darkness, ALAN/nnEMF, and geoengineering (e.g., aerosol-induced dimming) lower solar EMF (UV-A/B), which induces cellular hypoxia and blocks Becker’s regenerative current from the beginning. Without reestablishing cytochrome c oxidase to make DDW hydrate melanin, you never get to produce the one trillionth of one ampere of current needed to turn stem cells into de-differentiated pluripotential cells that can heal damage. Moreover, UV-A light stimulates NO, which controls mammals’ stem cell depots. So this is also turned off. Hypoxia-inducible factor-1 (HIF-1), stabilized under low oxygen or light stress, shifts metabolism to glycolysis (Warburg shift), increasing lactate production, and lactate use drives éR, which drives tissues into the Warburg redox shift. The pictures at the beginning of this blog are all there for you to review.
- HIF-1 and PER2 Link:
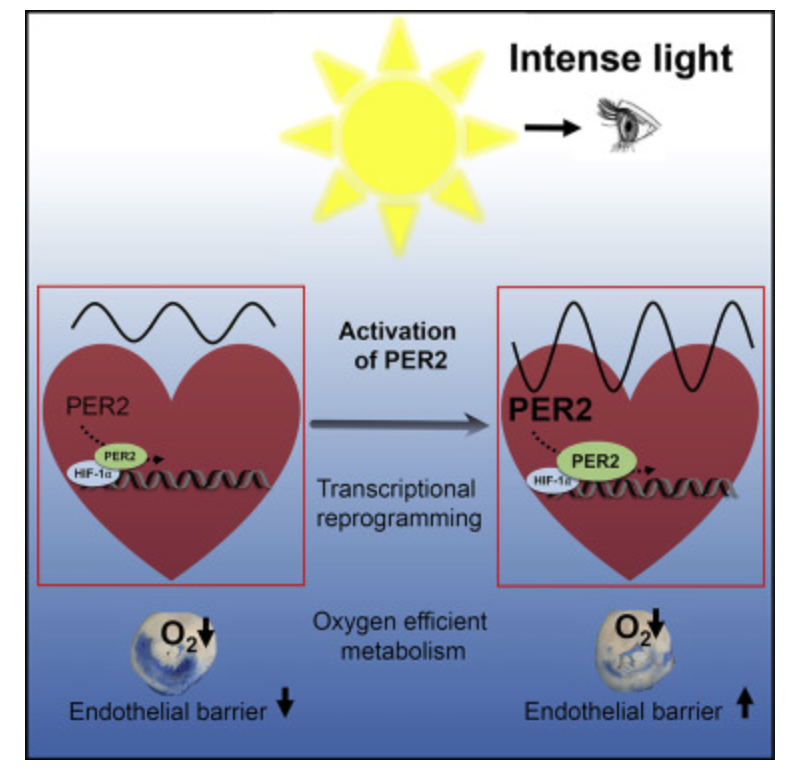
- HIF-1 belongs to the same protein family as Period 2 (PER2), a core circadian gene. Liu et al. (2012) show PER2-HIF-1α-dependent regulation of SIRT3 (sirtuin 3) under hypoxic conditions, modulating the choices between TCA cycle flux and glycolysis. Sunlight enhances PER2 expression, optimizing oxygen use, while ALAN/ darkness/geoengineering disrupts this, favoring HIF-1α and Warburg metabolism. If you read the links on my forum in the Mitochondrial Thermodynamics DIY, you’d already know all of this—->https://x.com/DrJackKruse/status/1613298172801044482
Red Light as a Therapeutic Equivalent: Tiina Kuru
- 43% of the sun is Red light (e.g., 600-1000 nm), and PBM/LLLT can mimic sunlight’s effect, upregulating PER2 and downregulating HIF-1α, reducing éR and Warburg metabolism. The red light was critical in the evolution of all heme-based proteins after the Cambrian Explosion. Tiina Karu’s work supports red light as a “drug” equivalent, enhancing mitochondrial cytochrome c oxidase and oxygen utilization, countering the cellular effects of dehydration and hypoxia in an oxygen-filled atmosphere.
- PER2-mediated ischemic preconditioning of the heart links sunlight/red light to optimal cardiac oxygen delivery, disrupted by mtDNA damage at cytochrome c oxidase (favoring Warburg shift).
- HIF-1 belongs to the same protein family as Period 2 (PER2), a core circadian gene. Liu et al. (2012) show PER2-HIF-1α-dependent regulation of SIRT3 (sirtuin 3) under hypoxic conditions, modulating the choices between TCA cycle flux and glycolysis. Sunlight enhances PER2 expression, optimizing oxygen use, while ALAN/ darkness/geoengineering disrupts this, favoring HIF-1α and Warburg metabolism. If you read the links on my forum in the Mitochondrial Thermodynamics DIY, you’d already know all of this—->https://x.com/DrJackKruse/status/1613298172801044482
- Historical and Evolutionary Context:
- The Great Oxygenation Event (driven by sunlight/photosynthesis) coupled oxygen, sunlight, and PER2, enabling mammalian survival. Dinosaur extinction (darkness, cold) reflects HIF-1 explosion in non-therapod dinosaurs, while mammals adapted via PER2. Modern sunlight avoidance (geoengineering, indoor living, atrophic skin, ALAN) increases cardiac/PAD death and hypoxia-related diseases. With sunlight from the KT event, trillions of photosynthetic algae could now grow uncontrollably to make oxygen, transforming the entire planet’s atmosphere and setting up the perfect storm for the evolution of a mammalian mitochondrial world post-KT until human technology changed the signal when we innovated ALAN.
Disease Implications:
- Blue Light/nnEMF and Vitamin A Liberation from opsins:
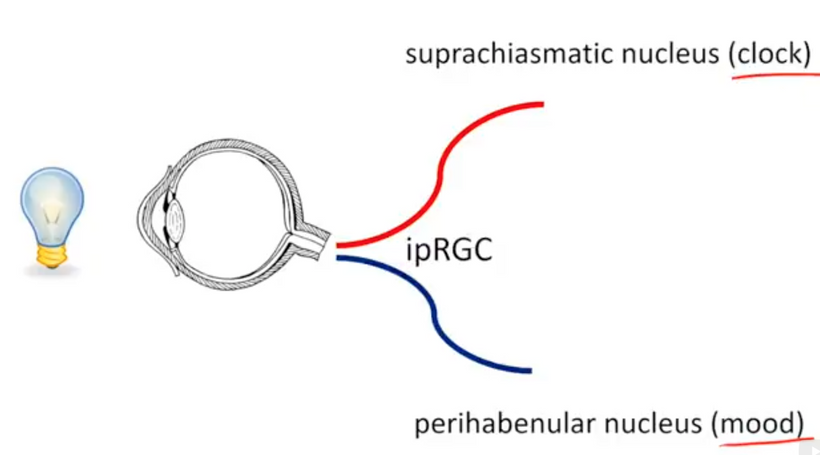
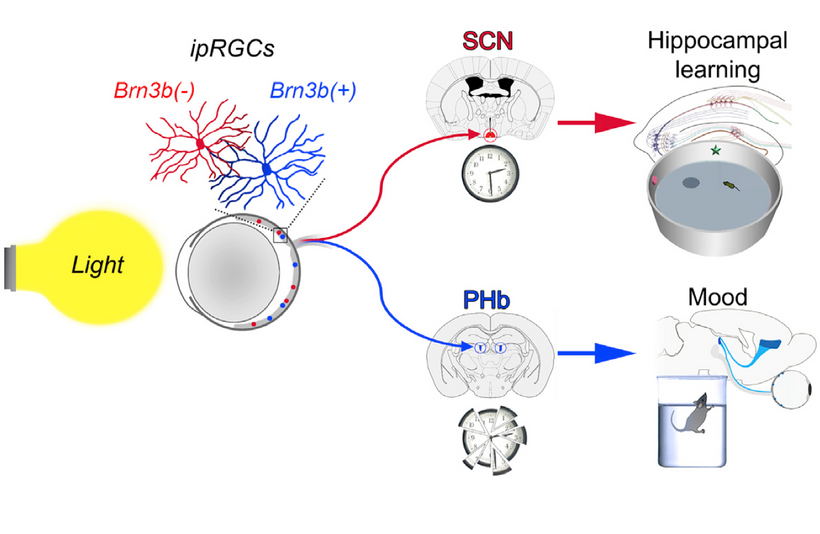
- Blue light and nnEMF liberate Vitamin A from cells and membranes, raising its presence in blood plasma as an aldehyde (retinaldehyde). This lowers plasma Vitamin C and D levels, disrupting local antioxidant defenses (melatonin/ISR) and circadian signaling. Retinaldehyde destroys small molecule modulators of the mammalian circadian mechanism, including PER1 and PER2, critical gears in the eye clock of the SCN. Rev Erb Beta and alpha are the destroyed nuclear circadian receptors that are also heme-based.
- Impact on Circadian Rhythmicity and mtDNA:
- Disrupted PER1/PER2 periodicity (from retinaldehyde toxicity) leads to quantum timing loss in mtDNA, accelerating mutations. Human mtDNA mutates 15–20 times faster than nuclear DNA (vs. 5–10 times in primates), and nnEMF accelerates this further, driving faster epigenetics and chronic disease creation. This is why MAHA is HAHA. They remain myopically focused on food when light is the driver of all chronic diseases.
- Cellular Disorganization and Energy Cost:
- mtDNA mutations of the heme-based proteins cause cellular disorganization by impairing information processing (energy = information in physics). Energy is trapped in cells’ electronic level, stored in vibrational/electronic bonds (e.g., PER/HIF-1), membranes, gradients, fields, and mtDNA-structured water. nnEMF increases ubiquitin marking, raising protein turnover, robbing energy, lowering redox state, and increasing éR, spreading heteroplasmy across organs and manifesting many new disease phenotypes.
- mtDNA Vulnerability:
- mtDNA is more vulnerable than nuclear DNA due to: (1) higher oxidative stress (mitochondria as ROS source), and (2) the matrix-side negative membrane potential concentrating lipophilic cations 1,000-fold, amplifying nnEMF effects. This charge density change mimics vasopressin’s effect on water (negative charges QE has a blog on it too!), linking light to metabolism and controlling all the biochemicals to control it (Light > Food).
Warburg Shift and Heteroplasmy:
- nnEMF increases epigenetic expression, raising mtDNA error rates and heteroplasmy (normally 10% per decade, accelerated by nnEMF). This energy-intensive process lowers the redox state, increases éR, and drives Warburg metabolism (increased lactate, glycolysis), fueling disease (e.g., ALS, cancer, diabetes, obesity).
- Warburg-like metabolism (increased glucose uptake, lactate) is a hallmark of cancer (FDG-PET), atherosclerosis, sarcoidosis (granulomas), and neurological conditions (e.g., schizophrenia, elevated lactate). The Ras-ERK-PI3K-mTOR axis, via mTORC1, stabilizes HIF-1α, promoting glycolysis (HK-II, GLUT1) independent of oxygen and driving disease progression.
- Compared to complete glucose oxidation, Warburg metabolism generates less ATP stochastically but creates a higher éR, which enables more rapid glycolysis (10–100 times faster) during the light stress event. This fuels macromolecular synthesis (e.g., via the pentose phosphate pathway [PPP] and fatty acid synthesis), supporting cell growth and division.
- The rapid shift to Warburg metabolism is triggered by changes in the organism’s light environment (e.g., nnEMF/blue light vs. solar EMF), a phenomenon misunderstood in biochemistry due to the neglect of biophysics and photo-bioelectric currents. Red light drops blood glucose and insulin, while blue light and nnEMF raise blood sugar and insulin levels. The pictures above state that case. Any nnEMF disrupts mitochondrial electron flow, increasing éR and lactate, while solar EMF (UV-A with IR) restores NAD⁺/DDW, lowering éR and supporting oxidative phosphorylation of the TCA cycle. This is why you need to see AM sunlight to use the TCA cycle. Every time the day’s first light is not the sun, you just Warburg shifted your LIFE toward disease.
- The Great Oxygenation Event (driven by sunlight/photosynthesis) coupled oxygen, sunlight, and PER2, enabling mammalian survival. Dinosaur extinction (darkness, cold) reflects HIF-1 explosion in non-therapod dinosaurs, while mammals adapted via PER2. Modern sunlight avoidance (geoengineering, indoor living, atrophic skin, ALAN) increases cardiac/PAD death and hypoxia-related diseases. With sunlight from the KT event, trillions of photosynthetic algae could now grow uncontrollably to make oxygen, transforming the entire planet’s atmosphere and setting up the perfect storm for the evolution of a mammalian mitochondrial world post-KT until human technology changed the signal when we innovated ALAN.
Elevated lactate and éR drive reductive stress, and this recycles NAD+ to NADH rapidly (high NADH/NAD⁺), signaling the release of vasopressin, which initiates the Integrated Stress Response (ISR) and this activates GDF15, which suppresses anabolism but supports rapid energy needs (e.g., cancer proliferation, retinal repair). This dual role explains pathological (e.g., AMD, cataracts, retinal damage, RP progression) and adaptive (e.g., retinal resilience) outcomes. ALAN turns on the VP-ISR-GDF15 axis while red light radiation seems to turn it off.
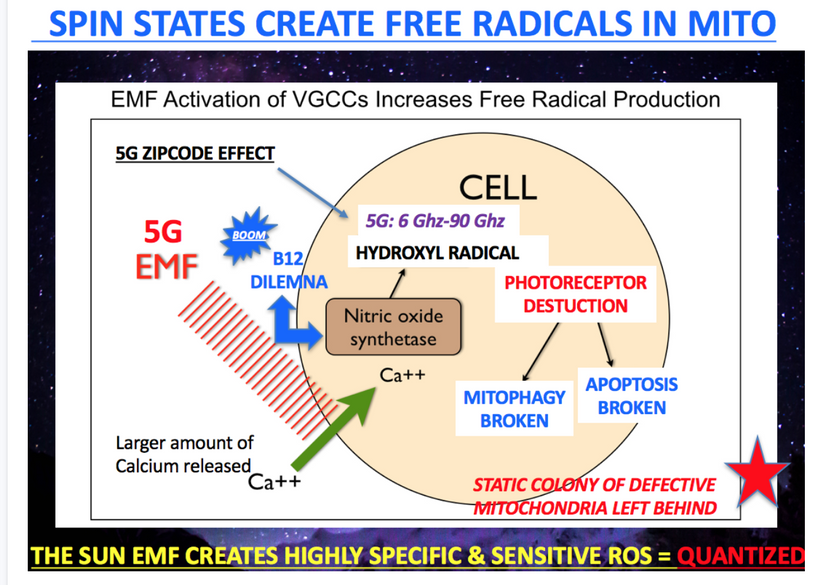
Prediction: High nnEMF exposure in Big City USA like NYC, intensified by 5G and ionospheric radiation, alters cellular pH (e.g., <7.35), driving a Warburg redox shift and contributing to the crash at an early age for humanity
- Mechanism: nnEMF activates VGCCs, increasing Ca²⁺ influx and ROS/RNS (peroxynitrite, ·HO), disrupting the NAD⁺/NADH ratio and lowering cytochrome c oxidase activity (pseudohypoxia = low NAD+). This reduces DDW production, dehydrates melanin, and increases NaCl concentration, dropping intracellular pH below 7.35 (-20 mV) and membrane potential toward zero mV. The night owl lifestyle and blue light exposure amplify free retinal toxicity, damaging photoreceptors (melanopsin, NO, Rev erb /alpha-beta, cytochromes), while endurance stress (exercise) and/or mold burden (low alpha-MSH) enhance glutamine addiction via GDH dysfunction. This “toxic barrel” shifts metabolism to glycolysis, causing mitochondrial failure (energy crash) and neuronal dysfunction (cognition crash).
- Vasopressin’s Role in ISR Activation: Vasopressin (VP), a hormone released by the hypothalamus in response to osmotic stress, dehydration, or nnEMF-induced cellular stress, activates the ISR by stimulating eIF2α phosphorylation via eIF2α kinases (e.g., GCN2, PERK). This global translational arrest induces stress-responsive genes (e.g., ATF4, CHOP) and upregulates GDF15 (growth differentiation factor 15), signaling energy resistance (éR) systemically. nnEMF and artificial light at night (ALAN) increase VP secretion by disrupting melanopsin signaling and inducing osmotic stress (e.g., via dehydrated melanin). VP activates ISR, enhancing GDF15 production, suppressing anabolism (e.g., protein synthesis in the damaged area blocking regeneration), and promoting catabolism, exacerbating chronic conditions under reductive stress (high NADH/NAD⁺, lactate).
- Outcome: Low pH (e.g., intracellular pH <7.3), high lactate (blood >2 mmol/L), and low NAD⁺/NADH ratio (<1), measurable via blood gas analysis and metabolomics.

2. Dehydration from nnEMF lowers éR and pH —–> https://osf.io/preprints/osf/hgnmj_v2
- Prediction: nnEMF-induced dehydration increases conductivity (high NaCl, low water), reducing éR and pH and preventing recovery post-crash.
- Mechanism: nnEMF dehydrates cells, raising NaCl concentration and lowering pH (e.g., <7.35), reducing the membrane potential (-90 mV) toward zero mV. Dehydrated melanin becomes conductive, amplifying ultraweak biophotons and ROS/RNS, disrupting Becker’s regenerative current. Free retinal toxicity from nnEMF/blue light damages catalase and cytochromes, impairing H₂O₂ breakdown and DDW production and exacerbating the redox Warburg shift (light).
- The patient’s lack of solar EMF (night owl, indoor living NYC life) prevents pH restoration, perpetuating mitochondrial stress. People forget that before eukaryotes, only two domains of light used the Warburg metabolism because of the light present. It had nothing to do with food. This defined the Cambrian explosion and KT events, as I laid out on the Huberman/Rubin Tetragrammaton podcast. String the lessons together and integrate them. No one is.
- Outcome: High serum osmolality (>300 mOsm/kg), low éR (microcurrent <1 µA), and persistent fatigue/cognition deficits, measurable via pH electrodes and imaging.
3. Warburg Shift Reflects Light Environment Choice, not a diet
- Prediction: The patient’s exposure to nnEMF/blue light (vs. solar EMF) altered the mitochondrial free radical blueprint, driving the Warburg shift and cognitive crash of people in a nnEMF environment.
- Mechanism: nnEMF (5G, blue light) and night owl behavior increase AMPK and ROS/RNS via VGCCs, lowering NAD⁺ and pH, shifting metabolism to glycolysis (Warburg light effect). Solar EMF (UV-A) supports quantized free radicals and redox potential, but its absence (due to indoor nnEMF dominance) disrupts cytochrome c, enhancing glutamine addiction. Free retinal toxicity damages photoreceptors, amplifying this shift, while mold exposures (low alpha-MSH) and endurance stress compound mtDNA heteroplasmy.
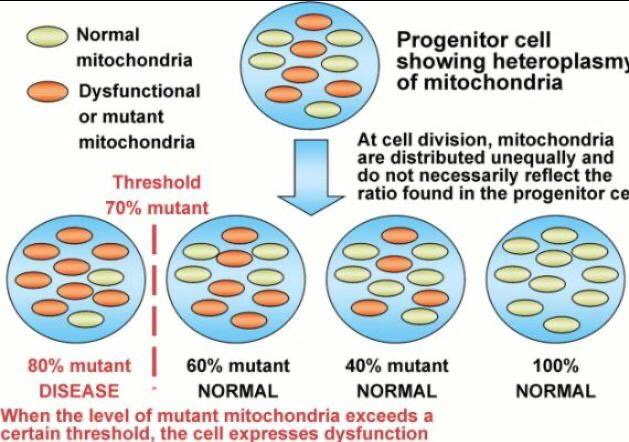
- Outcome: High AMPK activity, low cytochrome c activity (enzyme assays), and elevated glutamine/glutamate ratio, reversible with solar EMF.
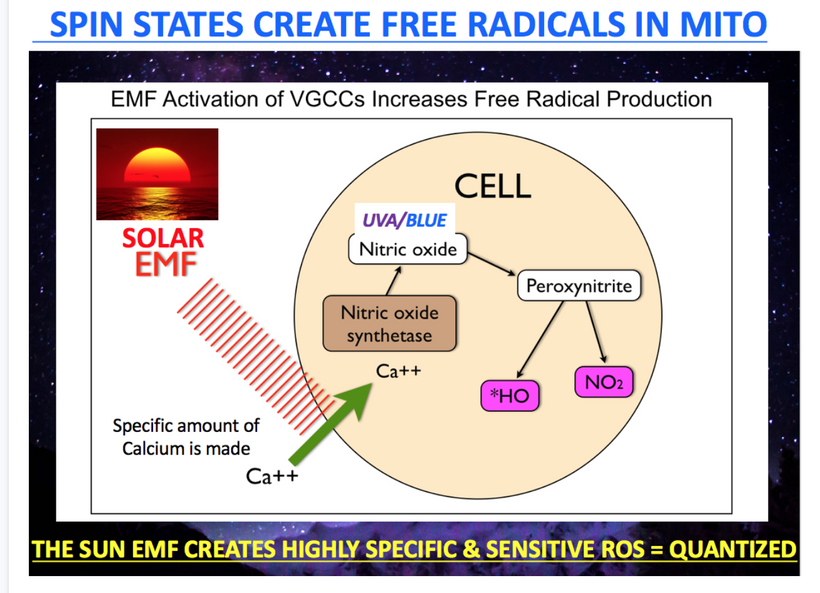
4. AM Solar EMF Restores pH and Redox Potential
- Prediction: AM solar exposure (30–120 minutes daily) restores cellular pH (7.35–7.45), reverses the Warburg shift, and improves energy/cognition.
- Mechanism: AM sunlight (UV-A, visible light) reduces free retinal, regenerates melanopsin/cytochromes, and boosts cytochrome c oxidase for DDW production, raising NAD⁺/NADH and pH to -20 to -25 mV. This hydrates melanin increases éR, and counters nnEMF-induced dehydration, restoring membrane potential (-90 mV) and bioelectric signaling. Solar EMF’s quantized free radicals support redox potential, mitigating the Warburg shift and mold effects.
- Outcome: pH normalization (7.35–7.45), improved NAD⁺/NADH (>2), energy (fatigue scale <10), and cognition (MoCA >28) within 8 weeks.
5. Comprehensive Redox Protocol Prevents Progression To FORMAL CHRONIC DISEASE
- Prediction: A redox-first protocol (AM sunlight, grounding, DDW, PBM, hypertonic saline, methylene blue) restores pH/éR, reverses cognitive crashes, and prevents neurodegeneration.
- Mechanism: AM sunlight and PBM (810 nm) regenerate photoreceptors/cytochromes, grounding reduces ROS/RNS, DDW enhances DDW production, hypertonic saline improves conductivity, and methylene blue repairs mtDNA, raising pH and éR. This requires an MD to implement who knows how to assess patients. This counters nnEMF/blue light, free retinal, and the Warburg shift, restoring mitochondrial and neuronal function. Detox (mercury, mold) must follow REDOX to clear the “toxic barrel,” preventing re-toxification.
- Outcome: Sustained pH (7.35–7.45), VCS score >7, energy/cognition recovery (MoCA >28, fatigue <10 at 6 months), and reduced Alzheimer’s risk (normal amyloid-beta on CSF).
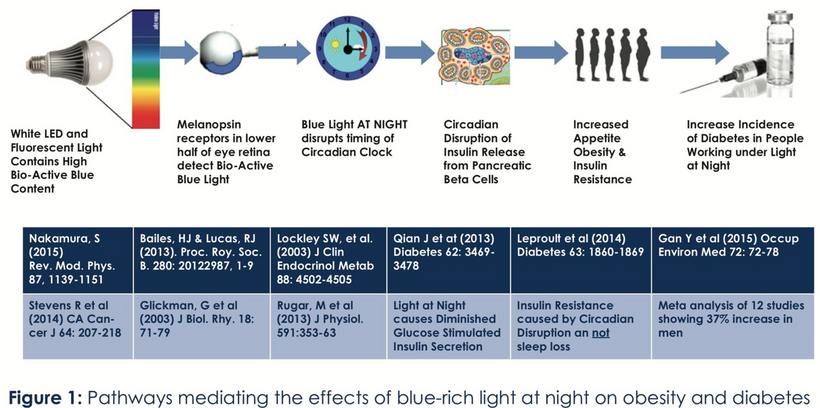
My decentralized interpretation of the Warburg effect is a marker of circadian mismatch, not a fuel source. Moreover, it is well supported by recent findings in the literature. The old idea that fuel source is critical is now on life support. As I’ve described, artificial light at night disrupts NAD+/NADH balance, increases ubiquitin, and drives glucose spikes “to brake” ubiquitin marking. AM UV and natural light cycles restore NAD+, lower ubiquitin, and normalize metabolism, reducing cancer, obesity, ALS, and autoimmunity, as I’ve emphasized with plants and animals sharing this response in the ubiquitin series on my website. The picture below lays it out AGAIN.
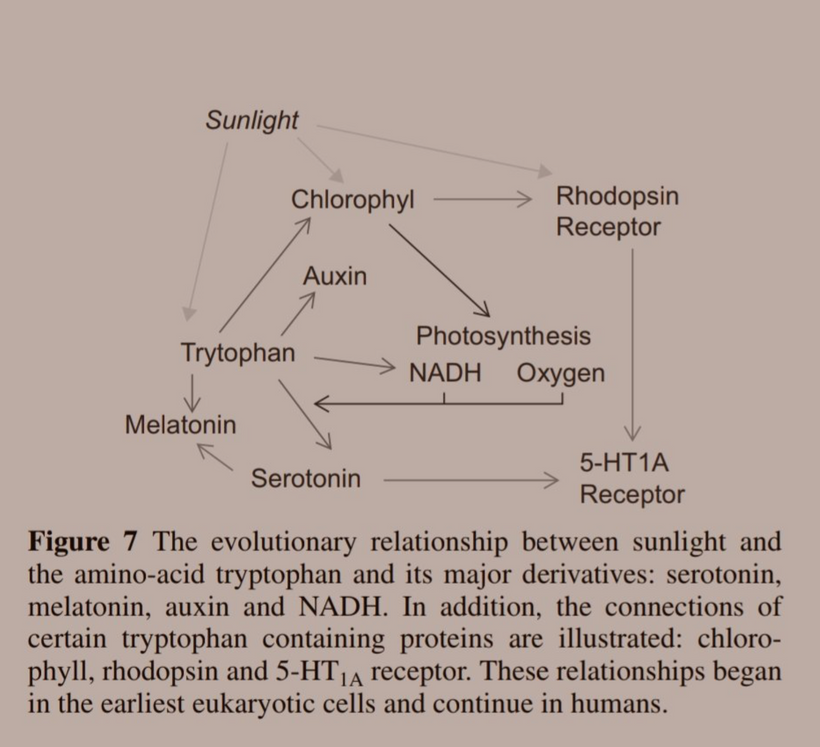
- All of these proteins above are heme-based and have tryptophan embedded in them. They explain why chlorophyll came first in evolution, heme proteins came second, and now we are at melanin today. All of this was tied to light variation, temperature changes and oxygen tension variation in the atmosphere of LIFE.
- Altered Mitochondrial Metabolism: Blue light/nnEMF oxidizes heme in cytochrome c oxidase (MT-CO1), stalling oxygen reduction to water. This leaves O₂ as ROS, which excites molecules (e.g., lipids, proteins) to emit biophotons. Warburg redox shift (light) controls metabolism photo-bioelectrically (glycolysis dominance) because it operates when the inner mitochondrial membrane loses delta psi. This increases ROS from incomplete ETC activity (e.g., Complex III leakage via MT-CYB), amplifying biophoton output. This is why mitochondrial damage always leads to more biophoton release, spreading the failure at the speed of light.

- Photo-Bioelectric Networks: Biophotons travel through cellular water, extracellular matrix, and bioelectric fields (e.g., gap junctions, membrane potentials), signaling mitochondrial distress. Popp suggested biophotons regulate cellular quantum coherence; in my model, their chaotic increase under stress disrupts this, spreading dysfunction. This is why the nuclear spins of atoms are lost. You need optimized optical pumping to maintain quantum coherence for health and longevity.

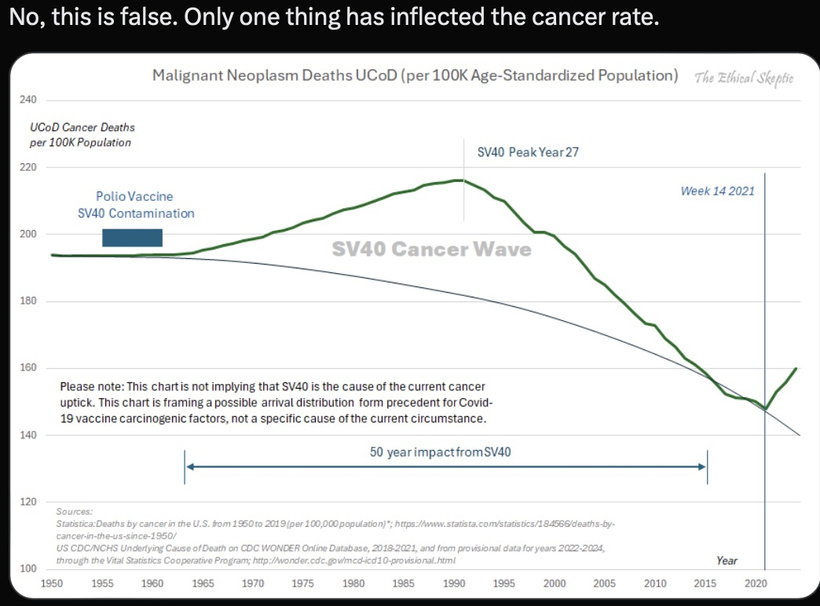
- Are there other mechanisms in jabs that explain rapid-onset dementia, cognitive decline, and prion-like changes? Could it be the source of the negative polarity in things that give a thing a net negative flux? Recall that all living things have a serious net negative charge. Also, recall that water goes from a neutral polarity in its bulk state and then gains its net negative charge in the structured state when UV and IR/NIR light excite water. This structured state excludes protons and creates coherent domains that act like electromagnetic capacitors for the crystalline lattice in cellular water that resists hydrophobicity. HYDROPHOBICITY IN medications with substantivity effects on water CAN allow for the possibility of REVERSE TRANSCRIPTION TO CAUSE NOVEL DISEASES. Did you know this? Slide the above points out. Jabs do this.
- Might the answer be that water and sunlight naturally create unbelievable amounts of electrons and protons free of an energy charge? Hydrophobicity can break this effect. This allows DNA to unwind and be copied under reverse transcription, which can cause turbo cancers. This effect would be magnified if SV40 contamination was present. DNA plasmid numbers increase the probability of such an event. Translation is widespread in annotated noncoding sequences, including untranslated regions (UTRs), introns, and long noncoding RNAs (lncRNAs), especially in cancer, aging, and neurodegeneration. 3’UTRs follow the mRNA coding sequence, regulate localization, stability, and translation, and control the selection of hydrophobic amino acids that change protein folding in cells.

Impact on Oxygen-Dependent ETC Proteins
The hardest-hit heme proteins—hemoglobin (Hb), myoglobin (Mb), Rev-erb, and ETC cytochromes—are very oxygen-dependent, making them prime targets for biophoton-mediated damage. Understanding how technology is making us ill is key.

SUMMARY
Because of the atomic organization of human cells (AMO physics), energy should always be available within the system, provided oxygen and water are hydrating our semiconductive heme and melanin proteins. When this relationship is altered for any reason, the disease manifests. The energy derived from the sun is stored coherently and ready for use over all space-time domains. Mitochondrial water production is critical in our mammalian blueprint.
The fidelity of this water creation is the basis of organisms’ autonomy and it was built around the GOE and the KT event in our history. As a result, modern organisms are never simply at the mercy of their environments because of the coherent energy stored. When the environment steals this ability from cells (nnEMF), cells are at the mercy of food and exercise.
More to the point, we don’t have to eat constantly (Leptin Rx), leaving plenty of time for other beneficial, pleasurable activities (SEX).
The other consequences are that the organism is exquisitely sensitive and free from the mechanical constraints of life on Earth and satisfies, at least, some of the basic conditions for quantum coherence. Water provides that as well.
Liquid water on Earth is quantum coherent even at ordinary temperatures and pressure. This is why Nature got the idea to build the IMM of cells around liquid water. It functionally is a naturally formed quantum computer.
Liquid water made in the mitochondrial matrix is the most wonderous chemical Nature has ever built because the water forms more coherent domains than the water from the hydrology cycle. Even latitude variation shows how water homogeneity changes as solar inclination affects its molecular arrangements and bond angles. Mitochondrial matrix water associates with macromolecules and membranes in cells into a gel-liquid crystalline configuration that enables enzymes and nucleic acids to function as quantum molecular machines that transform and transfer solar energy at close to 100% efficiency. Liquid crystalline water at interfaces also provides the excitation energy that enables it to split into hydrogen and oxygen in photosynthesis, simultaneously generating electricity for intercommunication and the redox chemistry that ultimately powers the entire biosphere on the 3rd rock from the sun.
Water is the means, medium, and message of life. For you to remain healthy, it must be made in large quantities in your mitochondrial matrix. Any reduction in its production will lead to disease.