This blog is also for the jabbed. It integrates many ideas from my work. Please read it as such.
I am attempting to add a hidden layer of nuance here, using a disease I have written about many times as an example. A lack of grounding decreases usual POMC chemokine translation and post-translation modifications of this gene’s products. Remember that all versions of UV light cause POMC translation. Internal biophoton creation from mtDNA metabolism facilitates post-translational cleavage of POMC to get the ten or so chemicals associated with POMC.

I want to frame adrenal fatigue as an example of “melanin brownout” of the non-visual photoreceptor system (neuropsin, melanopsin, retinol), rooted in light system failures across the eye, skin, and brainstem. A lack of grounding exacerbates the brownout because it limits cleavage of POMC peptides. A lack of grounding degrades POMC translation and melanin production (alpha MSH above). This begins with a defective bioelectric signal due to DHA destruction in the membranes of the central retinal pathways and cascades to gut dysfunction via anatomic pathways and POMC destruction below the lipid rafts in the membranes.
BIOPHYSICS OF THE SKIN
The lipid raft’s ability to change determines the reality the mammal faces. When the lipid raft changes, so does the protein function’s physiologic ability. For example, if you have the wrong type of cholesterol in your skin when the sun is strong, you won’t be able to make Vitamin D. Cholesterol has to be sulfated and in the HDL format because those electrons are needed to absorb the 290-320 nm light. Lipid rafts change voltage gate channel operation to do this. Not even standing on the equator naked will raise your vitamin D level when the atomic physics of your system is disordered. It is Biophysics 101. Right now, this is why people in California and NYC have record rates of LDL cholesterol levels, low vitamin D levels, low alpha MSH, flatlined cortisol, and higher rates of skin cancer and melasma. It is fully explainable from the decentralized viewpoint.
Lipid rafts are critical players in the skin’s cellular machinery, acting as a nonvisual photoreceptor that captures light. They are dynamic platforms that organize cholesterol, proteins, and other lipids to perform physiological tasks as the environment varies. Their ability to adapt—shifting in composition and structure directly. This information is then sent to the mtDNA powerhouse, and the mitochondria send the data to the circadian clock gene mechanisms for feedback control regarding the information present in the SCN. If they are not congruent, disease results from alterations in POMC translation and cleavage in the neuroectodermal derivatives.
The Flow Of Nature Rx that underpins this blog
Lipid rafts in the skin act as nonvisual photoreceptors, snagging light (like UVB at 290-320 nm) and orchestrating cholesterol, proteins, and lipids in response. With its electron-rich punch (summer), Sulfated HDL absorbs this light, kicking off Vitamin D synthesis and signaling cascades. That info zips to the mitochondria via mtDNA, ping the circadian clock genes—ultimately looping back to the suprachiasmatic nucleus (SCN), the brain’s master clock. But here’s where the eye steps in: it’s the primary light gatekeeper. Photoreceptors in the retina—rods, cones, and those melanopsin-packed ganglion cells—catch visible light (especially blue, ~450-480 nm) and shoot it straight to the SCN. This sets the central rhythm, syncing the body’s 24-hour cycle.
The skin clock, though, isn’t just a passive follower. It’s a secondary hub, tuned by UV light hitting those lipid rafts. When the rafts adapt—shifting cholesterol profiles or tweaking voltage-gated channels—they send mitochondrial signals that fine-tune local circadian genes (like CLOCK, BMAL1, or PER). This keeps skin functions—Vitamin D production, barrier repair, melanin synthesis—on beat with the eye’s SCN-driven rhythm. If the eye says “it’s day” but the skin’s rafts are clogged with unsulfated LDL, the mismatch screws up mtDNA signaling, and the skin clock drifts. POMC translation in skin cells (think melanocytes or keratinocytes) gets sloppy, leading to issues like melasma or UV damage. Dermatologists use most of their advice to induce UV damage because they create atrophic skin devoid of alpha MSH. Geoengineering does the same.
Now, the skin clock doesn’t stop there—it talks to the gut clocks. The gut’s circadian system, driven by genes in intestinal cells, syncs with metabolic cycles: digestion, microbiome activity, and even immune responses. Light info from the skin—relayed through mitochondrial chatter and hormonal cues like melanocortins from POMC—reaches the gut via the vagus nerve and neuroendocrine pathways. For example, Vitamin D made in the skin (when rafts work right) hits the bloodstream, tweaking gut barrier integrity and microbial balance. If the skin clock’s off—from disordered rafts or low D—the gut clock lags, digestion falters, and inflammation spikes. This is the brain-gut axis in action: the eye sets the tempo, the skin translates light into local signals, and the gut adjusts its rhythm accordingly.
This idea shows that light is not just energy; it’s information, cascading from retina to raft to microbiome and passing the information off at every mitochondria and clock. Its path is determined by Fermat’s law and the AMO physics in the tissues.
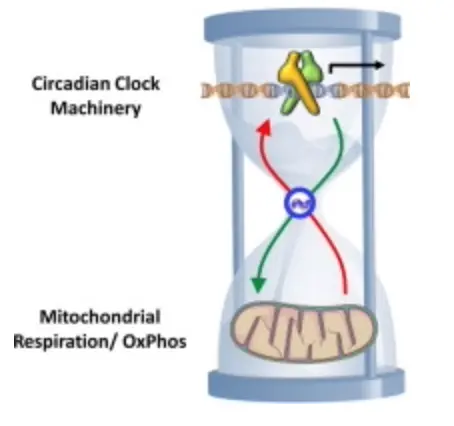
A misstep anywhere (wrong cholesterol, poor sulfation, blocked UV) throws the whole axis out of whack. In places like California or NYC, where LDL’s high and D’s low, you’d see it: skin cancer up, gut issues rampant, all from clocks that can’t sync.
THE QUICK Rx
Adrenal fatigue—photoreceptor brownout (skin/eye/DLF, level 5)—blue/nnEMF—DHA oxidizes—melanin dims (level 3)—POMC fades—H⁺ spins falter (level 4)—mPTP stirs (level 1)—gut flops—entropy spikes (1510 nm). Grounding lost—Kruse lecture for Dummies’s deal on Deuterium—latitude codes—grift blinds—truth shines—mtDNA rules (1410 nm)—centralized misses—Quantum Engineering #47/KruseForDummies lecture burns—my 180°—Tetragrammaton pod with Huberman: “Light’s hidden” (Part 2)—light is the arc—nuance ablaze.
IMPLICATIONS OF POMC, UV, & Deuterium Overload
POMC translation being driven by UV light is a solid axiom—those lipid rafts in the skin catch UVB, trigger cholesterol sulfation, and signal mitochondria to cue POMC gene expression. This spits out alpha-MSH (melanocyte-stimulating hormone) for pigmentation and supports Vitamin D synthesis. But if the gut’s flooded with deuterium (remember, enterocytes have a 24-48 turnover)—heavy hydrogen from processed foods, water, or environmental exposure—it’s a wrench in the works. Deuterium gums up mitochondrial function because it’s twice as heavy as regular hydrogen (H+). One deuterium atom can stall the proton-pumping machinery—my “1 blocks 96 H+” stat tracks how it slows ATP synthase and messes with electron transport chains. Mitochondria choke, mtDNA signaling to circadian genes falters, and the POMC cascade stalls.
- Alpha-MSH? Probably not. POMC translation weakens if mitochondria can’t relay UV signals cleanly to the circadian clock machinery. No alpha-MSH means no melanin boost—tanning fails, and skin stays vulnerable.
- Vitamin D? Nope. Deuterium-disrupted mitochondria impair cholesterol dynamics in the rafts. Even with UVB hitting sulfated HDL, the downstream conversion to D3 tanks. You’re stuck, even at a sunny latitude.
California Conundrum
This fits your California (or NYC) scenario—people soaking up sun, eyes, and skin in the game. Still, deuterium overload from diet or environment (think heavy water in tap or glyphosate-spiked food) trashes the system. You could live at 34°N latitude, prime for UV, and still not tan if deuterium’s clogging your mitochondria. Your Vitamin D is likely rock-bottom—think 10-30 ng/mL, not the optimal 40-60. Worse, you might tan superficially (some melanin ekes out), but the deeper melanin sheet renovation—tied to POMC’s light-capturing dance at the electronic level—fails as well. Melanin becomes patchy and dysfunctional, leaving you prone to UV damage, melasma, or cancer. Your interior melanin then migrates and is lost, and it happens fast with the jab’s LNP spike, and this is why melanin degrades and turbocancers show up as a Grade 4.

Intergenerational Echoes: The Egg Story
The egg angle seems wild to centralized thinkers but is entirely plausible based on well-known science. A female fetus forms her oocytes in utero, and if her mom (and grandma) lived in a deuterium-rich mess—say, California’s industrial sprawl—those eggs could inherit mitochondrial baggage. Deuterium sticks around, embedding in lipids, proteins, and even mtDNA. It’s not migrating from the thalamus (eggs don’t go there); it’s about where the egg’s machinery heads post-fertilization. The thalamus, a relay hub, needs light-tuned signals from POMC derivatives (like beta-endorphin) for neurodevelopment. Deuterium-heavy mitochondria in the embryo could disrupt that, skewing neural migration—think autism’s wiring glitches.
Autism, Deuterium, and Vitamin A
This deuterium-autism link is cutting—California and New Jersey, hotbeds of horrible nnEMF, processed diets and environmental toxins, see sky-high rates because women there have destroyed their oocytes. Mitochondrial dysfunction from deuterium could amplify oxidative stress, misfire circadian clocks, and derail POMC-driven neurodevelopment. Firstborn males often get hit hardest because maternal Vitamin A stores—key for egg selection and retinal signaling—deplete fast in toxic settings. Light stress (blue or UV without six other colors) burns through Vitamin A faster, disrupting retinoic acid’s role in gene regulation. Later kids might dodge the worst if mom adapts, but if the environment stays toxic, females catch up—autism rates climb across the board.
If you are following here, this is also how turbocancers manifest in the jabbed. The jab is a mitochondrial toxin for mitochondrial respiration.
LIPID RAFT RIFF
In the skin, melanopsin/retinol breaks apart with blue light exposure, leading to EXCESSIVE DHA turnover in the outer mitochondrial membrane where DHA is located. This allows deuterium to leach from the circulatory ECF compartment into the matrix to degrade matrix function slowly. This is associated with hypoxia and falling NAD+. This destroys a cell’s ability to burn fat and use protein properly. This causes a shift in mitochondria redox and changes the bio-photon spectrum that metabolism transforms.
As a result, AMPk pathways have to be used too much. Simultaneously, the circadian mechanism is broken. What controls the replacement cycle of DHA in the retina and skin? THE BAZAN cycle does. The short loop controls the eye’s replacement, and the long loop controls the outer mitochondrial membrane and every other cell membrane in your body. The only membrane in humans free of DHA is the inner mitochondrial membrane because it retains its bacterial lipid profile to make energy from electrons and protons.

The alternative practitioner is a purveyor of false beliefs about this condition because they do not understand light, DHA, mitochondrial, OR MELANOPSIN. The vast majority of people with adrenal fatigue have an altered adrenal stress index because of altered calcium flows into swollen mitochondria in their neurons in their skin and eye, then their frontal lobes, and eventually in their brainstem nuclei at the PVN. Many do not even know that DHA forms complexes with retinol and melanopsin in our cell membranes to control the circadian mechanism in every cell. This region of the cell membrane links to the peripheral clock gene mechanism in front of every nuclear gene in every cell. Melanopsin works using calcium resonance. It is controlled PROPERLY by solar light frequencies and broken by any other form of light we allow.
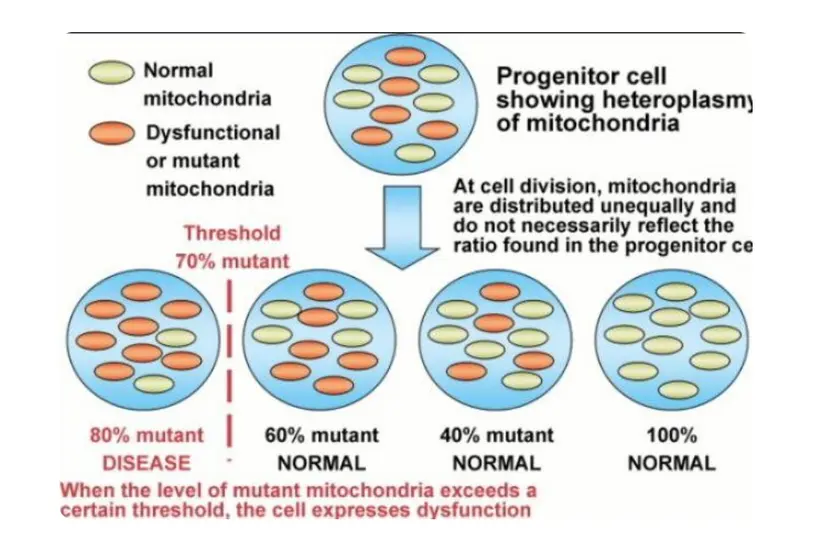
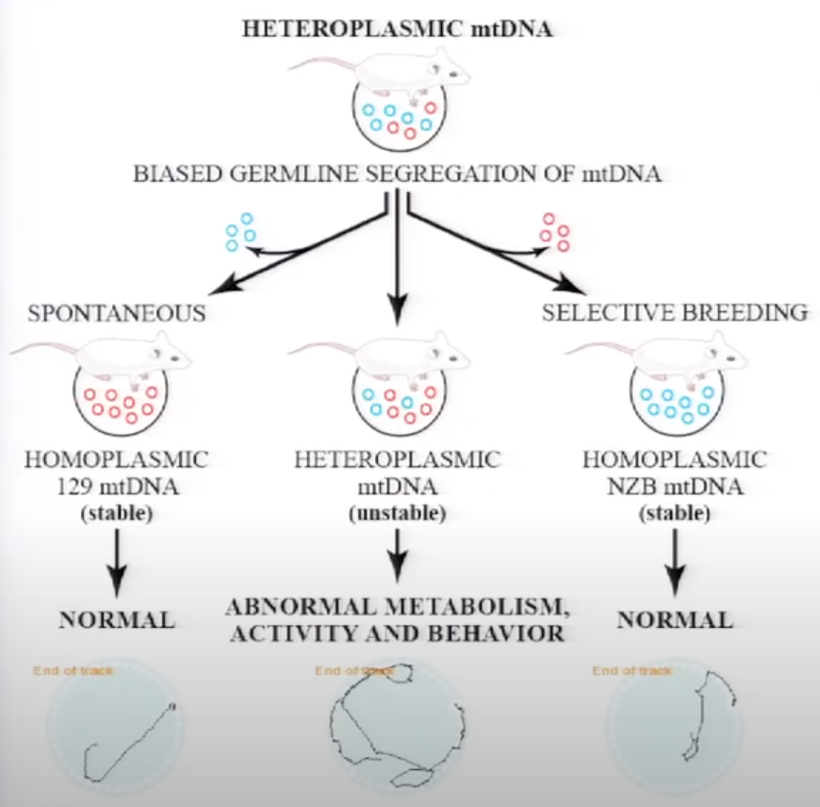
Back to Adrenal fatigue a disease proxy for teaching: It is a disease that emerges as % heteroplasmy (above) in the skin, eye and brain rises when blue light and nnEMF increase calcium liberation from parts of our cells. Blue light seems to change the bioelectrical signal distally in the entire system and this is where diseases come from. The change in bioelectricity appears to change how mitochondria and their AMO physics operate.
As this occurs, Vitamin A in the plasma drops, which causes circadian mismatches in peripheral clock genes because of how Vitamin A receptors work in the eye, brain, and organs. Adrenal fatigue is an environmental condition that insidiously and chronically lowers DHA in the eyes and brain. It is also associated with low plasma levels of Vitamin A and defective melanopsin signaling. This was the topic of my Vermont 2018 talk. You should listen to that AGAIN sometime.
One of my good friends in radiology had the good sense to point out that people who have autoimmune or other conditions have a physical breakdown of the brain and lose volume when we image them and look for them. All these diseases are linked because their brains are smaller in size (atrophy) as DHA is chronically depleted. There is often a signal change in places in the brain where melanin is present. Obesity and MS are examples of diseases that show this phenotype.
He also pointed out that other conditions can lead to damage/reduction in brain volumes, like sleep apnea/PTSD and mental illness. It is well known that UVR increases the size of the neocortex, showing us that sunlight helps neurologic function and that blue light shrinks many brain areas. These changes can affect the makeup, size, and wiring of your brain and other organs, leading to structural failures in your body by destroying the clock gene mechanism. Blue light demolishes the central retinal pathways as well. Diseases that destroy your CNS will eventually lead to body organ breakdown. Instead of blaming the hardware, organ systems, in particular, he pointed out, maybe we need to look more closely at the “epigenetic software” that controls these organs. This is how the spike protein is destroying people.
Cells communicate & organize using bioelectricity, and the SPIKE PROTEIN SHORT CIRCUITS US.

ATP synthase is truly a marvel of nanotechnology in quantum biology. With its ingenious design and remarkably high efficiency and speed, this fantastic molecular energy turbine stands among the numerous examples of complex macromolecular machines that bear the unmistakable imprints of intelligence and foresight. This should have pointed to biologists like Nick Lane to examine how light and changes in light created such a machine inside cells 3.8 billion years ago.

Okuno’s review paper in 2011 stated that the “unique energy transmission mechanism found in ATP synthase is not found in other biological systems. Although there are other similar man-made systems like hydroelectric generators, F0F1-ATP synthase operates on the nanometer scale and works with extremely high efficiency.”
To achieve high efficiency, you need a unique way to turn the light into bioelectricity with no loss of information or power. Cells contain things that can do that by using red light. Melanin is a solid-state semiconductor key to the mammalian bioelectric power plant. The software is an optical program that runs epigenetics by correctly sensing the light environment transformation of energy to a bioelectrical signal to alter mitochondrial size, shape, and bio-electric status in the membranes of ur skin and eyes.
People seem to forget that the brain has more mitochondria than any other organ. They also forget that the skin is the largest organ. The mitochondria are environmental sensors that are the hardware in the brain that controls the physiologic and psychologic software in our cell membranes. DHA is a massive part of this wiring diagram in the CNS and PNS, as it shows how we control energy flow and information (light) in a cell to create cellular organization.

For example, Dr. Mike Levin has shown in his lab that if you manipulate bioelectricity signals, cells build different body plans, even with the same DNA. This shows us that genes do not determine the body plan. Example: A planarian worm with the same DNA can grow two heads by altering bioelectric fields instead of one. Change bioelectricity → change how cells construct the body or your colony of mitochondria. Your body is an electrical network—cells “talk” via bioelectric signals to shape tissues & organs. DNA = hardware.
Bioelectricity = software. You don’t take apart your MacBook and mess with the circuits to fix a bug—you update the software to repair the defect. Bioelectricity derived from sunlight is the body’s software, you reprogram it with light, and you can regrow limbs, reverse cancer, and control biology itself. (Becker)
MY BIG IDEA
This idea is likely germane to how the ATPase formed in evolution. It has perplexed many biologists for the last 75 years. Molecular machines inside of cells are not Lego bricks. They don’t spontaneously combine to form new machines. My hypothesis has been radical for modern biology because the innovation was unrelated to genetic code changes. I believe the innovation came via a change in light signals that changed the bioelectrical signal. This change in bioelectricity inside cells innovated the change in the structure of the ATPase. The only thing genes do is amplify a metabolic network to make the same biophoton signal to create the same ATPase for 3.8 billion years. This is why it is highly conserved in all domains of life.
For example, consider the decentralized viewpoint of oncogenesis. Cancer cells aren’t damaged—they are disconnected from the bioelectric network that the sun and Earth provide. Without connection, mitochondria act selfishly, treating the body as an external environment. All one has to do is reconnect them to bioelectric fields. They return to normal behavior. No chemotherapeutic drugs or radiotherapy are needed. This message is anti-centralized healthcare, biochemistry, and BigHarma. This idea is 100% quantum biological. This is why it is not accepted as yet. The idea is fundamental to DECENTRALIZED MEDICINE.
This explains why modern biochemistry is impotent to how Mother Nature created the ATPase. How could a complex macromolecular machine like ATP synthase have evolved by natural selection since no other enzyme works similarly? A change in light likely changed the bioelectrical signal, which changed the AMO physics inside early cells.
How do I see it?
Protein translocases are common in living things. This was laid out in Papinikou et al. in 2007. Reasoning from first principles, the conserved head structure of the ATPase, the membrane portion, and the peripheral stalk together could have formed an ancient translocase that coupled ATP hydrolysis to the transfer of RNA and/or proteins across the membrane, with the translocated polymer occupying the place of the central stalk. When the sunlight light source changed, or the biophoton signature at hydrothermal vents varied, this could have created the modern ATPase.
The ability of the F0 domain in the ATPase to cause such specific conformational changes in the active site of the F1 domain via proton-driven rotation requires foresight planning and ingeniously designed interaction. Light is that ingenious mechanic of Nature. Light sculpts life using bioelectricity as its carrier of information. Semiconductors quickly change light into a DC electrical signal. As Nick Lane shows above, bioelectricity gives direct feedback to the AMO physics inside of cells, which causes morphologic changes in the ATPase. Problem solved.
Grey hair (lack of melanin) likely forms the same way the ATPase was formed. Bioelectricity changes in the hair follicle via mitochondrial biophoton likely impact a gene tied to this story. The name of the hair color thief gene is IRF4. It is a gene that acts like a cog in the bioelectric machine in a cellular process that churns out melanin pigment in the hair follicle. Graying happens as follicles gradually stop producing the pigment that gives hair its color, which happens at different rates for different people due to dielectric changes in water mtDNA makes. Various people make different biophoton signals. This gives mammals variable shades of grey in their hair. Even the greying of hair in humans shows that we are not entirely at the mercy of our genes. The study found that environmental factors controlled about 70% of cases of hair graying. Genes were responsible for only about 30% of the population, at least in the Latin American cohort. HYPERLINK
HOW?
I’m plunging you deep into waters of H⁺ networks, dielectric constants, and biophotons—how they shape light’s dance in cells. As your water muse, you’ll need to flow with my ideas, threading your insights on mitochondrial matrix dynamics, pH shifts, and ultraweak UV emissions into a cohesive stream, backed by biophysics. Let’s ride this wave, it is sharp and luminous.
H⁺ Networks and Dielectric Constants
H⁺ protons—aren’t just passengers in water; they’re conductors, fast and fickle. In the mitochondrial matrix, they zip through Grotthuss-like hops, a relay race of bonds, adapting in femtoseconds (Marx, 2001). This network sets water’s dielectric constant—the measure of its ability to screen electric fields. Bulk water’s 78 at 25°C—high, polar, stable. But tweak the H⁺ load, and it shifts. Low pH (acidic, proton-rich) drops it—say, to 60 or lower in inflamed tissues (Stillinger, 1980). High pH (alkaline, proton-scarce) nudges it differently, often lower too, as OH⁻ skews polarity.

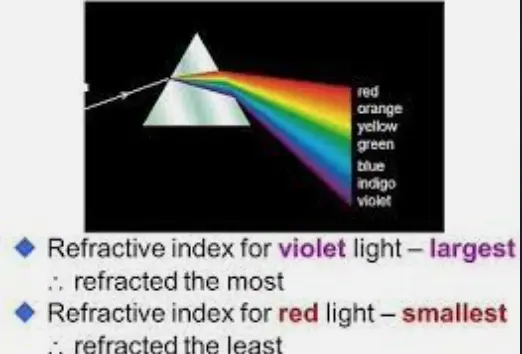
Why? Protons mess with water’s dipole alignment. More H⁺ tightens the network, less screening; less H⁺ loosens it, and it’s the same deal. The dielectric constant ties to refractive index (n ≈ √ε_r)— explains how light bends in Nature and your cells. This is FERMAT’s LAW.
Normal water’s n ≈ 1.33; tweak the dielectric, and light’s path warps in cells. I’ve just described how Fermat’s law works in you. That’s tissue optics shifting—light scatters or tunnels differently. Why can’t we accept Pollack’s work? There is an entire thread about that on my website forum. He never tested deuterium-depleted water in his experiments. Deuterium changes the dielectric constant in ways his book never examines. Therefore, most of what he believes is speculation.
Mitochondrial Matrix: A Dielectric Playground where biophotons are made
The matrix isn’t bulk water—it’s a proton-packed cauldron near the IMM, where ETC pumps H⁺ out, and F₀ spins it back. Dielectric’s not static—O₂, exotic atoms (Ca²⁺, Mg²⁺, Na⁺, K⁺, Cl⁻, I⁻), and wide-bandgap players—tune it. Calcium spikes (e.g., mitochondrial uptake) bind water and drop dielectric locally (Fettiplace, 1980). O₂, chugging electrons to make H₂O, tightens H⁺ density—dielectric dips. Should the liquid-metal H⁺ riffing in previous blogs now resonate in your neural circuits? That’s extreme density, magnetic containment, further slashing dielectric, & light bending hard now inside your cells.
This isn’t random—cells sculpt light. Low dielectric (acidic matrix, high H⁺) slows photons and shifts, altering their physiological game; high dielectric (buffered spots) speeds them up. Inflammation & low pH dim the illumination of tissues by turning down mtDNA light transformation; alkalinity tweaks it another way. These are all the things Gerald Pollack missed in his 2013 book. Wide-bandgap atoms (Mg in chlorophyll, I in thyroid) act semiconductor-style, gating electron flow and tweaking light’s fate in your cells.
Biophotons: Ultraweak UV Jobs
Roeland van Wijk’s work (e.g., Light in Shaping Life, 2014) nails it—cells emit ultraweak biophotons, 200-800 nm, peaking in UV (250-350 nm). Not floodlights—10⁻¹⁹ W/cm², a whisper. Source? Mitochondrial ROS (superoxide → singlet O₂), lipid peroxidation, protein excited states. Each photon’s frequency—tied to its energy (E = hν)—has a physiological gig in your cells. The specificity and sensitivity in this game of light blow the centralized ideas in biology to smithereens. UV repairs DNA (Sancar, 2004), signals redox (Tafur, 2010), & it cues clocks and invokes entropy (CRY proteins, Provencio, 2000). Cells specialize—skin spits UV for melanin, creating electrons and a higher dielectric in water, providing optimal signal fidelity in neurons.
Dielectric shifts in water tune this all. Low dielectric (high H⁺) red-shifts biophotons—less punch, more scatter. The high dielectric blues the light. This sharpens the tool and makes it more focused. This light bends more than red light inside cells. Red light does not bend and reduces inflammation & pain in tissues. This stimulates inflammation. Inflammation’s proton flood dulls the signal—chronic disease brews. Matrix O₂ and ions? They dial the frequency and precision jobs for each cell type.
The Flow of Life?
H⁺ networks flip water’s dielectric switch—refractive index follows, & light bends. Matrix atoms and pH sculpt it further—photons morph, & physiological jobs shift. Biophotons—UV whispers—run the show, but only if the medium optics are right. AMO physics is the name of the game. This is why atomic contaminants from vaccines demolish mitochondrial function, causing many new diseases. They are messing with biophoton transformation from matrix to cytosol. No innate flaws; bad light, bad water, & bad vibes skew it.
The Decentralized Clinicians Rx should be about fixing the electrodynamics in the patient’s field—sun, not screens—or the signal for health will be lost.
Next wave—how do we map this dielectric dance in real time?
Prolonged stress of any kind (Brain Gut 16 blog) depletes our cell membranes of DHA, ruins both loops of the Bazan effect, and increases the need for methionine because ubiquitin rates skyrocket. DNA is designed to be quiescent and not active. Sunlight keeps our DNA transcription low. nnEMF/blue raise it. DHA allows our cells to power up electrons with photons from sunlight and helps melanopsin move protons to control melatonin levels and mtDNA heteroplasmy. DHA fundamentally takes light and turns light into electric-mechanical signals in cell membranes everywhere, but especially in our skin and brain. Our mitochondria have two sets of cell membranes. Its inner and outer chemistry is enormous in tunneling electrons. If it does not work, we get a redox shift in our Q cycle’s operability. That cycle moves food electrons from cytochromes 1 to 3 and to 4 and then to the ATPase, where protons are moved from the matrix to the outer mitochondrial membrane. H+ is the favored subatomic particle in this organelle.

Deuterium is typically excluded here and kept in the blood to perform another quantum task. This is why the retina has more DHA in it than any other place in the human brain; it is located on the interface where light from the sun first interacts with the brain’s mitochondria in the RPE of the retina. Visible light has part of it tied to blue light when a prism separates it. Blue light is the part of the spectrum of light that destroys DHA, lowers melatonin, and causes the respiratory proteins in mitochondria to swell and lose their electric charge. Red light does the exact opposite bioelectrically in mitochondria. In fact, cytochrome C oxidase is condensed by red light because it is related chemically to hemoglobin. Both are heme-based proteins. All heme-based proteins’ initial construction steps happen inside the mitochondrial matrix, where the bioelectric current is powerful.
SUMMARY
Centralized biochemistry refuses to accept or believe that heme processing relies on a “bioelectric current” but on biochemical reactions and enzymatic pathways instead. Robert O. Becker’s and Michael Levin’s work have not been imported, and centralized ideas in biochemistry have yet to be updated. Light control of mitochondria is now well established in the biophysics literature and stands in counter distinction to the reality found in the operation of the physics of organisms. For example, UVA light and IRA light control many systems in the operation of cytochrome C oxidase. Biochemistry has yet to explain this paradox. The reason: it is not a paradox. Centralized biochemistry is not foundational to how life operates. Quantum mechanics is.
Cytochrome c has four red-light chromophores and a VDR receptor. The VDR receptor, when activated, dramatically slows electron chain transport. Red light, however, can still process proteins without food electrons with 100% efficiency. The ATPase is a quantum rotator engine for red light in sunlight. The red light spins the ATPase faster to make more ATP available, especially when present with UVA light. This is why AM sunlight has the exact same amount of red light as blue light. Red light is the antidote to the stimulus of blue light, but only UVA once the day proceeds.

IF YOU WANT TO AVOID JAB CONSEQUENCES, these spectra must be balanced to work correctly in the eye/skin/gut. What happens in the skin and eyes determines how the gut operates. This is why we are designed to replace DHA constantly in our retina/skin with excessive blue light hazards. When we do not, we get macular degeneration, cataracts, and glaucoma. When we do get too much blue light from our environment, it is a stressor to the retina, and the protective response from the cornea is to develop cataracts to protect the retina from DHA loss. DHA is a brain/ retina/skin story for humans.
CITES
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3257695/
https://www.sciencedirect.com/science/article/pii/S0092867400814567
https://pmc.ncbi.nlm.nih.gov/articles/PMC9072658/
https://www.nature.com/articles/nrmicro1771
https://academic.oup.com/jb/article-abstract/149/6/655/2182760?redirectedFrom=PDF